
摘要
背景
体细胞胚胎发生(somatic embryogenesis, SE)提供了许多商业机会,其中最重要的是能够无性繁殖具有稀有或优良性状的单株。对于许多长寿的物种来说尤其如此,比如针叶树,但它们的长一代时间带来了实质性的挑战,包括随着植物年龄的增长,对SE的抵抗力增加。鉴定了一个体细胞胚源性树的克隆系,该克隆系的茎原基对SE诱导反应持续了十多年,为研究成年白云杉茎组织中SE的分子基础提供了一个独特的机会。
结果
在SE诱导一周后,利用微阵列分析对取自该应答基因型的茎外植体进行转录组全表达谱分析,与非应答基因型的外植体进行比较,鉴定出每个基因型中四个差异最大的表达基因。使用绝对qPCR将分析扩展到诱导后三周,结果显示所有八个候选基因的差异表达一直保持到诱导处理结束,尽管程度不同。最引人注目的是,候选基因在无应答基因型中的表达量和持续时间都表明了强烈的生理反应。通过对它们假定身份的检验,进一步揭示了这四种蛋白质编码与已知在生物防御中发挥重要作用的被子植物蛋白质相似,并且它们在长时间内的高水平诱导与生物防御反应的激活是一致的。相比之下,响应型基因型中更温和的反应,包括针叶树特异性脱氢酶的诱导,更符合适应性应激反应的引发。
结论
虽然需要更多的证据来确定SE反应性与特定生理反应之间的关联,但这些结果表明,生物防御激活可能是拮抗的,可能与它引发的大量转录和代谢重编程有关。未来工作的一个主要问题将是确定如何以及是否可以使用抑制生物防御激活来促进更有利于SE诱导的生理状态。
背景
植物体细胞胚胎发生(SE)已经成为园艺、农业和林业工业中的一种成熟的生物技术,为商业规模生产无性系幼苗提供了能力[1- - - - - -3.]。然而,诱导胚性组织形成(SE诱导)的效率仍然存在问题,特别是在木本物种,如针叶树中。例如,虽然合子胚胎属于少数物种松科科是高度敏感的,许多其他针叶树种要么完全不敏感,要么生产效率太低,无法在商业上可行。一个更突出的问题是成树组织的顽固性,如果克服这种顽固性,就可以无限制地繁殖具有优良特征的单株树[4]。
虽然对诱导培养基的明智操作已经成功地提高了合子胚胎的SE诱导效率,特别是对松树[3.],到目前为止,成功应用于营养组织的案例最多也只是少数[4,5]。此外,尽管被子植物中有许多影响SE诱导的生理和遗传因素[6- - - - - -8然而,由于缺乏有效的实验系统,即使是确定针叶树营养组织中SE诱导的最基本方面的努力也受到了阻碍。
为了解决这一缺陷,十多年前开始的实验以体细胞胚胎衍生的白云杉为目标,期望它们比种子生长的树木具有更大的SE诱导倾向。这导致了白云杉无性系的鉴定,该无性系产生的芽即使在达到性成熟后仍对SE诱导有反应[9]。结合针叶树基因组学的进展[10- - - - - -12],这为探索SE诱导在成树云杉芽原基中的分子方面提供了前所未有的机会。
利用最近构建的针叶树32k寡核苷酸探针微阵列[12],转录组全表达谱鉴定了其中四个差异表达最多的基因,并在诱导第7天鉴定了一个无反应基因型。使用绝对qPCR将分析扩展到第21天,发现这些候选基因的表达动态存在实质性差异。最明显的是,在无应答基因型中,候选基因表达的幅度和持续时间都更大,这表明对诱导治疗有强烈的生理反应,可能对SE诱导有拮抗作用。对其假定身份的检查进一步揭示了这种强烈的反应可能是生物防御引发的结果,而反应性基因型的中度反应暗示了适应性反应。
结果
原始芽内体细胞胚胎发生的诱导
在应答基因型(G6)的原始茎外植体中,SE诱导的详细描述已经被描述[9]。简单地说,先对芽进行消毒,切除原芽切段,然后置于SE诱导培养基上(图2)1A)。由于期望差异基因表达可能与SE诱导的响应性(或缺乏响应性)有关,在诱导一周后,用从外植体中提取的RNA进行了微阵列分析(图2)1B).这个时间点的选择是基于经验观察,这表明它足够早,可以避免胚胎团块形成产生的偏差,这种偏差可能会混淆与SE诱导相关的基因的鉴定,而不是那些在胚胎发生过程中变得活跃的基因。在诱导的前两周,G6和无应答基因型(G12)的外植体都表现出针状原基的伸长和在针状原基的切割面和基部形成少量愈伤组织的特征。在诱导的第三周,一些G6外植体在细长的针状原基上或愈伤组织内产生结节,并伴有微量的胚胎团块(EM),这标志着SE的开始(图2)1C)。在诱导的第四周,部分G6外植体产生了快速增殖的EM(图2)116周后,480个G6茎部外植体中有22个(4.6%)产生EM,而480个G12外植体中没有一个产生EM。

白云杉成树原始枝的SE诱导。(一)纵切的前花蕾代表了SE诱导收集的花蕾。(B)诱导处理一周后的茎原基外植体,收集组织进行微阵列分析。(C)胚团(EM)的形成发生在一些G6外植体中,大约在诱导后三周。(D)随后的增殖产生EM,然后可以继代培养并用于产生无限数量的幼苗,这些幼苗是收集芽的亲本树的克隆。D:诱导处理天数,棒材= 0.8 mm。
微阵列分析和候选基因的选择
微阵列分析使用收集点(第0天)和SE诱导一周后(第7天)的外植体进行,每个时间点对每种基因型分析5个生物重复(详见方法)。第0天和第7天的基因型内差异是实质性的,G12和G6中分别有4381和5807个靶点存在差异表达(图1)2A)。虽然这包括共享的3602个靶标,但差异表达靶标的总数(6586)代表了微阵列上所代表的23,854个不同的白云杉基因的相当大的比例[12]。基因型间比较揭示了许多微小的差异,167个靶点在第7天有显著差异,而第0天有27个靶点(图2)2B)。比较基因型内折叠差异的大小进一步支持了对诱导治疗反应的相似性(附加文件)1).这种适度的差异表明,诱导治疗在涉及的基因方面产生了很大程度上的共同反应。然而,这并没有考虑到表达水平的数量差异,这是在qPCR分析中检查的一个方面(见下文)。

基因型之间的差异表达(DE)和对SE诱导处理一周的反应。(一)7天SE诱导处理产生的基因内DE(学生t检验;校正p值<0.05)。(B)诱导治疗前后基因型间DE靶点(学生t检验;校正p值<0.05)。(C)DE与基因型、治疗和相互作用效应的关系基于双向方差分析(p值< 0.05)。没有根据折叠差的大小进行过滤,所有统计分析都进行了多重检验的校正。
为了进一步研究基因型和诱导处理是如何相互作用的,我们进行了双向方差分析(图2)2这表明8433个靶点在所有组合中都存在差异表达,其中约90%仅与SE诱导处理相关。此外,两种基因型之间约有10%的差异,约3%的基因型与治疗之间存在显著的相互作用。值得注意的是,所有表现出基因型效应的靶标也对SE诱导有反应,其中37%表现出X基因型治疗相互作用。
为了确定qPCR分析的候选基因,根据诱导第7天相对于其他基因型的最大折叠差异对微阵列数据进行排序(附加文件)1),结果显示靶基因数量和差异基因表达量的变化趋势相似(表2)1).为了在第7天选择每个基因型分化最大的4个候选基因,详细检查了差异表达最多的靶点。结果显示,G6中排名前30位的基因中有9个属于一个小基因家族,该基因家族编码针叶树特有的脱氢酶DHN1的三个变体(图1)3.一个;额外的文件2),这是以前在挪威云杉中发现的[13]。由于它们的高度相似性,这些DHN1基因被认为代表一个单一的目标(表1)2).在剩余的3个G6候选基因中,只有一个与拟南芥(Arabidopsis)的apoplastic III类过氧化物酶AtPrx52高度相似3.).最后两个候选基因都编码了不寻常的蛋白质,这些蛋白质似乎是针叶树特有的,分别含有富含苏氨酸-谷氨酰胺和脯氨酸的重复片段(图2)3.B, C)。

五个最不寻常的候选基因的氨基酸序列比对。(一)DHN1是一种针叶树特有的脱氢酶,被发现由三种变体组成(见附加文件)2获取更多信息)。保守的取代用黄色突出显示,并且两个脱氢结构域是固定的。(B, C)QT-repeat和富含脯氨酸的候选序列似乎是针叶树独有的,它们排列在一起,突出了它们高度重复的结构。(D)PgPI20a/b与具有代表性的被子植物同源物一致,其编码高度保守的小分子量丝氨酸蛋白酶抑制剂,属于MEROPS I20家族(MER201390)的未分配亚类。杨树trichocarpa(EEF00358),拟南芥(At1G72060),玉米(EF406275)。参见附加文件3.剩余三个候选基因的氨基酸序列比对。
为G12的所有4位候选人找到了假定的身份(表1)3.).研究发现,前两个序列编码密切相关的蛋白质,与一种罕见的丝氨酸蛋白酶抑制剂(在被子植物中高度保守)具有高度的序列相似性,并根据SignalP 4.0分析预测具有氨基末端信号肽[14)(图3.D)。这些蛋白酶抑制剂的另一个显著特征是存在8个保守的半胱氨酸残基,这些残基符合CRP5550类富含半胱氨酸的小肽,这是一个非常大的排泄肽家族,包括防御素,以及许多其他抗菌蛋白[15]。与G6 PgPrx52一样,第三个G12候选基因编码一种外胞体III类过氧化物酶,但与拟南芥AtPrx21最相似3.).剩余的G12候选基因编码一种与拟南芥AtcwINV1最相似的细胞壁转化酶(附加文件)3.).
关于随时间的表达变化,将第7天与第0天进行比较,发现除了一个候选基因外,两种基因型中的所有候选基因都显着增加,而对应基因型中没有基因表达减少的例子。这表明,第7天的差异表达是由于原始基因型中较高水平的激活所致(表2)2和3.).同样值得注意的是,G12候选基因在G12外植体中的表达水平比G6候选基因在G6外植体中的表达水平更高,这表明无应答的G12基因型的一个主要区别特征是候选基因激活水平更高。
芯片分析与绝对qPCR的一致性
进行绝对定量分析大大提高了分析的分辨率,除了允许在多个样品内和样品之间直接比较任何基因的表达外,还可以直接比较任何基因的表达。这是通过我们小组开发的一种称为LRE qPCR的方法完成的,该方法极大地简化了绝对定量,很大程度上是由于无需构建目标特异性标准曲线[16- - - - - -18]。
如方法部分所述,9个内参基因的表达分析显示,在微阵列分析中使用的每个时间点进行的5个生物重复中,平均方差约为±20%(样本组内方差),这在一定程度上表明了LRE qPCR可以达到的分析精度[17]。此外,当比较四个样本组的平均表达水平时,9个参考文献中有6个样本组间方差低于±20%,反映了该实验系统中非常低的生物变异性水平。这反过来又避免了进行内参基因归一化的需要,而在基于qpcr的基因表达分析中,通常需要进行内参基因归一化,特别是对于那些采用相对定量的基因表达分析[19- - - - - -22]。
表格4总结了8个候选基因的表达水平,揭示了两种基因型中所有候选基因在第0天的表达都很低,每10 ng RNA的表达量在5.6到534之间。尽管如此小的数据集的生物学观点必然是有限的,但这确实表明所有候选基因在芽收集时都是相对静止的。此外,与微阵列分析预测的结果一致,G12外植体内G12候选基因的激活率平均比G6外植体内G6候选基因的激活率高9倍。
表格5比较两种基因型在第7天候选基因表达的相对差异,这为微阵列分析提供了广泛的证实。例如,微阵列和qPCR定量预测的基于折叠差异大小的相对排序,在两组候选基因内部和之间是基本一致的。一个明显的例外是qpcr衍生的QT-repeat候选序列的比值为340倍。然而,这是由于G12外植体在第7天的表达水平非常低(表1)4),使其与微阵列分析的可比性受到怀疑。这些数据集也反映了有限的生物学视角,只能通过分析两个时间点来实现。因此,本研究的一个主要方面是利用LRE qPCR的高容量将分析扩展到诱导的第21天。
候选基因表达动态分析
通过包括三个额外的时间点(3,15和21天)将分析扩展到第21天,可以更详细地定义候选基因表达的动态。例如,在G6外植体中,QT-repeat候选表达在第3天达到接近最高水平,这一水平一直维持到第21天,而在G12外植体中,其表达在整个诱导处理过程中几乎不存在(图2)4A)。在G6外植体中也发现了广泛的胞外过氧化物酶PgPrx52的差异表达,在第3天达到最大表达,但在第7天下降了3倍,维持到第21天的水平(图2)4B)。

G6候选基因在SE诱导21天的表达谱分析。每个点表示平均数量,以每10 ng RNA转录本的数量表示,标准差以条形表示。个别定量在附加文件中提供7。
DHN1表达的动态性质与PgPrx52相似,在第7天达到峰值,随后在G6外植体中第21天逐渐减少3倍,表明早期的、短暂的激活(图2)4C)。然而,DHN1的表达不仅在G12外植体中很明显,而且在第15天逐渐增加,这表明该G6候选基因的激活不具有基因型特异性。虽然富含脯氨酸的候选基因的差异表达维持到第21天,但两种基因型产生了相似的表达动态,再次反映了适度的(如果有的话)基因型特异性(图2)4D)。
对于G12,所有四个候选基因都表现出高水平的差异表达(图2)5).此外,在诱导处理期间,除PgcwINV1外,所有候选基因的表达均逐渐增加,均达到最高水平,平均比G6外植体中G6候选基因的最高表达量高约20倍(图2)4).总体而言,G12外植体内的表达动态与诱导处理的强烈和持续反应一致。

G12候选基因在SE诱导21天的表达谱分析。每个点表示平均数量,以每10 ng RNA转录本的数量表示,标准差以条形表示。个别定量在附加文件中提供7。
总之,qPCR表达谱证实了在诱导第7天基于微阵列分析的候选基因选择的有效性,因为在整个诱导处理过程中,所有候选基因在其原始基因型内保持差异表达,尽管程度不同。此外,总体生物学变异性足够低,足以揭示基因表达动态的强烈趋势,并说明绝对量化的效用,在其他属性中,提供了评估单个基因表达幅度的能力。与微阵列分析预测的结果一致的是,G12外植体中所有四种G12候选体的诱导水平都很高,并在第7天之后保持良好。这表明这些无反应外植体的一个主要区别特征是对SE诱导处理有强烈的生理反应。
讨论
植物外植体对许多类型的组织培养操作(包括SE诱导)的抗性长期以来一直是具有精英特征的单株克隆繁殖的障碍,这种能力具有重大的商业意义。这些还包括一些罕见的性状,如抗虫或抗病原体,这些性状的快速繁殖和传播可能具有重要的生态意义。这与森林树木特别相关,它们的长一代时间对传统的繁殖方法(如插枝生根)和基于传统育种的遗传改良计划构成了实质性的挑战。许多优良性状在成熟植物中变得最为明显,这一事实进一步加剧了这些限制,而在成熟植物中,许多木本物种,包括针叶树,对组织培养的操作变得不适应[4]。因此,鉴定出一株对SE诱导有反应的成年白云杉无性系,为解决SE诱导的分子方面提供了一个独特的机会,并期望这可能广泛适用于其他植物物种。
应激反应可能是SE诱导反应的决定因素
与se诱导研究中经常出现的情况一样,本研究的一个主要假设是,在反应性G6芽外植体中选择性表达的基因将成为研究的主要目标。事实上,表达谱显示了一个不寻常的谷氨酰胺-苏氨酸重复蛋白(图2)3.B),在整个SE诱导处理过程中特异性表达(图2)4A).尽管如此显著的差异表达暗示在诱导反应中有潜在的重要作用,但该蛋白似乎是针叶树特异性的,没有已知的功能,排除了该蛋白在体细胞胚胎发生中起普遍作用的可能性。富含脯氨酸的蛋白质也是如此3.C),直到第21天,G6芽外植体中表现出边缘性的差异表达,尽管持续存在(图2)4D);然而,这种蛋白质似乎是针叶树特有的,没有已知的功能。尽管剩下的两种候选基因编码具有假定功能的蛋白质(脱氢酶和过氧化物酶);数字3.A和附加文件3.),最大的差异表达发生在诱导处理的早期阶段(图2)4B, C),早在胚胎性组织开始出现之前,这再次不能很大程度上支持在决定SE诱导反应性方面的作用,至少不能直接支持。
与这种温和的反应相反,在无反应的G12芽外植体中,强烈的候选基因激活不仅持续到诱导处理的后期,而且达到了非常高的水平(每10 ng RNA最多有60,000-600,000个转录本)5).虽然绝对定量的有限使用迄今为止为普遍比较基因表达水平提供了不充分的背景,但EF1α确实为候选基因表达的大小提供了一些视角。作为内参基因,EF1α平均每10 ng RNA表达约700,000个转录本(见方法),这是本研究中遇到的表达最高的基因。根据EST克隆频率和微阵列分析(数据未显示),这与它是针叶树幼芽发育中表达最高的基因之一是一致的。因此,候选基因在无反应的G12外植体中的表达量表明,原始茎外植体的生理反应,特别是在SE诱导的早期阶段,可以决定其对SE诱导产生反应的能力。
虽然胁迫生理学的主题领域是广阔的,但很明显,被子植物描述的一些一般原理可以为针叶树芽外植体的生理反应提供见解,特别是与非生物和生物防御反应有关。例如,在外植体被引入培养后,预计会立即发生短暂的低强度氧化爆发,这表明在非生物应激(包括伤害)和生物防御反应中产生的早期氧化信号[23- - - - - -25]。然而,一个主要的区别是,生物防御反应的引发与第二次长时间的高强度氧化爆发有关,这反过来又引发了大量的转录和代谢重编程,包括防御蛋白表达的高水平诱导,生长的减缓,在最极端的情况下,诱导细胞死亡[23,24,26- - - - - -29]。这就产生了一个问题,即G12外植体产生的强烈反应是否反映了生物防御反应,使它们在生理上对SE诱导产生抗性,而G6外植体产生的适应性应激反应则产生了有利于SE诱导的生理状态。
尽管这种概括提供了一个有吸引力的模型,但应该强调的是,被子植物的防御反应已经被发现是一个高度动态和复杂的过程,涉及由水杨酸盐、茉莉酸盐和乙烯调控的信号网络之间的串扰,以及与其他植物激素的结合[27- - - - - -32]。因此,在没有直接支持证据的情况下,很难得出与针叶树芽外植体的具体相似之处。然而,通过检测四种G12候选基因(一种外胞体III类过氧化物酶、一种细胞壁转化酶和两种密切相关的细胞外丝氨酸蛋白酶抑制剂)编码的蛋白质的推测功能,支持了SE诱导处理在这些无反应的外植体中引发生物防御反应的观点。
外胞体III类过氧化物酶在生物防御激活中起着重要作用
III类过氧化物酶已被发现可产生外质体过氧化氢,作为生物防御触发的信号[27,33- - - - - -36]。最近有报道直接证明了这一点,其中玉米美国maydis(玉米黑穗病)叶片侵染可诱导单一III类过氧化物酶基因(POX12)的表达,该病原体的毒力依赖于其过氧化物酶抑制剂肽(Pep 1)的分泌[37,38]。此外,即使在没有Pep 1的情况下,诱导的痘12基因沉默也能恢复毒力,这直接证明了POX12的活性对于成功地建立针对这种病原体的生物防御反应至关重要[38]。G12外植体中III类过氧化物酶的持续、高水平激活(图2)5A)因此与生物防御诱导的作用是一致的。
另外,尽管是暂时的,支持这一论点的是,这个G12候选物在氨基酸序列上与拟南芥III类过氧化物酶AtPrx21最相似3.),它属于植物过氧化物酶的一个不寻常的进化分支[39]。损伤和微生物攻击产生的AtPrx21表达增加,这表明它对病原体具有保护作用[36]。AtPrx21在拟南芥中的过表达会产生抗性,这一事实更直接地证明了AtPrx21在生物防御中的作用葡萄孢菌(40]。因此,目前可用的数据表明,外胞体III类过氧化物酶基因的持续高水平表达在生物防御激活中起核心作用,G12 PgPrx21可能在针叶树中发挥作用。
相反,G6 III类过氧化物酶PgPrx52基因激活的短暂性(图2)4B)可能反映了适应性应激反应,其中初始氧化爆发在诱导治疗的前几天消散,恢复细胞氧化还原稳态[41- - - - - -43]。这与G6脱氢酶DHN1基因的激活相似,但稍微晚一些4C),这是一种针叶树特有的脱水剂,据报道在挪威云杉的芽休眠和越冬中起作用[13]。这也支持了G6外植体引发适应性应激反应的论点,因为脱水物质长期以来一直被认为在适应环境应激中起着基本作用。44]。因此,尽管是推测性的,但可以认为适应性应激反应可能是建立SE诱导反应的重要决定因素。
细胞壁转化酶的诱导
细胞壁转化酶的激活,在外质体内催化蔗糖水解为葡萄糖和果糖,也被发现在生物防御中发挥着重要作用,它提供了进行强烈代谢反应所需的大量能量[45- - - - - -49]。RNAi抑制表达为细胞壁转化酶在烟草防御反应引发中的核心作用提供了直接支持[50]。事实上,细胞壁转化酶诱导的突出作用导致有人建议将其归类为一种发病相关蛋白[45],这是一大类防御蛋白,在生物防御激活后高度表达[51]。
与拟南芥的氨基酸序列比较发现,G12转化酶与AtcwINV1(也称为at β frut1)最为相似(附加文件)3.),它是拟南芥中一个小基因家族的成员[47]。许多研究报道了拟南芥叶片病原菌感染诱导AtcwINV1表达[52- - - - - -54],除了农杆菌感染拟南芥细胞培养物后的诱导外[55]。重要的是,AtcwINV1在损伤后的选择性诱导也表明了在适应性应激反应中的普遍作用[56]。这与在G6外植体中观察到的PgcwINV1表达的逐渐增加是一致的。然而,与此形成对比的是,G12外植体在第3天至第7天之间的快速激活,平均达到G6外植体的两倍左右,这一水平一直保持到诱导处理的第21天(图2)5B).这可能反映了G12外植体在诱导处理的早期阶段有更强烈的代谢反应,这一假设与观察到的其余两个G12候选基因的高水平表达一致。
两种I20丝氨酸蛋白酶抑制剂的激活
两个高表达的G12候选基因被发现编码密切相关的蛋白酶抑制剂,属于MEROPS I20丝氨酸蛋白酶抑制剂家族的一个未分配亚类[57,58]。该亚类具有许多显著特征,包括存在一种瞬时肽,一种异常小的成熟蛋白(少于60个氨基酸),以及存在8种高度保守的半胱氨酸(图2)3.事实上,这些特征导致它们被归类为富含半胱氨酸的小肽超家族(CRP),这是一个非常大的分泌肽家族,由几百个基因组成。最初是基于与防御素的结构相似性,防御素是一种古老的抗菌肽[59],大量的crp已被证明在生物防御中发挥作用[15]。这类I20蛋白酶抑制剂的另一个显著特征是在被子植物中存在密切相关的同源物,主要是作为一个单一基因[60]。事实上,这种高度的保护导致Hartl等人。[60表明它们除了可能在生物防御中起作用外,还可能在植物生理学中起重要作用。针叶树中两个高度保守的同源物的存在为这种可能性提供了支持。
被子植物同源物与生物防御激活的直接联系来自于基因的表达分析美国maydisΔpep1-infected玉米叶片,如前所述,这引发了大规模的生物防御反应。在被诱导的220个基因中,玉米I20同源基因(EF406275)见图3.D)的诱导程度第四高,接种后24 h内增加了166倍(附加文件)1:表S1于[37])。同样地,拟南芥I20同源基因(At1G72060)是感染后第五大诱导基因木霉属harzianum, 24小时内增长了2.79倍[61]。将生物防御激发子应用于10日龄拟南芥幼苗中也可诱导该I20基因,细菌鞭毛蛋白肽Flg22在1 h和3 h后分别增加5.08倍和7.45倍(附加文件)1:表S1于[62])。在玉米叶片中也发现了玉米同源物(serpin)的氧化胁迫诱导作用,在施氢16 h后分别增加了2.91倍和7.56倍2O2和甲基紫紫素[63]。然而,拟南芥在非生物防御中的作用不太可能,因为转基因幼苗中拟南芥同源基因的异位表达显著降低了对氧化、渗透和盐胁迫的抗性[64]。
因此,两个针叶树I20同源物在G12外植体中的诱导动力学与被子植物中观察到的生物防御激活一致,到第7天,这两个基因每10 ng RNA接近20万个转录本,与第0天相比,大约增加了2000倍。另一个值得注意的特征是在整个诱导处理过程中表达逐渐增加,到第21天达到每10 ng RNA约600,000个转录本(图2)5因此,虽然这类不寻常的丝氨酸蛋白酶抑制剂的确切生化功能仍有待确定,但高水平的氨基酸序列保守性,结合它们的表达动态,为这两种针叶树蛋白酶抑制剂在针叶树生物防御中发挥作用的假设提供了支持,类似于在被子植物中观察到的。
向生理过程的范式转变可能对抗SE诱导
虽然人们早就认识到外植体的生理状态可能是对SE诱导的反应性的主要决定因素,但对其潜在机制知之甚少。此外,历史上绝大多数的研究工作都集中在确定促进SE诱导的因素上,例如,通过激活SE促进基因。这项研究扩展了这一范式,表明外植体的无反应性不一定是由于先天缺乏SE促进活性,但生物防御激活可能是潜在的主要拮抗剂。
生物防御引发的一些生理方面为这种假设提供了普遍的支持。例如,通过外源应用各种激发子激活生物防御已被证明可以显着降低植物生长,这是将代谢能量从生长重定向到防御的结果[65,66]。这包括茉莉酸盐(JAs)的作用,它是与伤害相关的生物防御激发的中心调节因子,已被证明直接介导从生长到产生生物杀灭化合物、细胞壁重塑和防御蛋白表达的转换[28]。事实上,JAs的生长抑制与细胞周期进程的抑制直接相关[67],除了直接拮抗赤霉素酸的促生长活性外[31],因此提供了证据,证明JAs可能是SE诱导的直接拮抗剂,部分基于细胞分裂是胚胎性组织形成所必需的假设。
另一种支持证据,尽管是间接的,来自蛋白质组学研究,该研究报告了生物防御蛋白(主要是致病相关蛋白)的表达与培养组织缺乏胚胎发生能力之间的相关性[68- - - - - -71]。与蛋白质组学分析的定量性质相关的另一个值得注意的观察结果是,这些非胚胎性组织中防御蛋白表达的幅度,通常是分析中最突出的蛋白质。虽然很难进行直接比较,但这与G12外植体中G12候选基因的强烈激活是一致的,并且假设将代谢资源转向生产如此大量的防御蛋白本身可能对胚胎性组织的形成是拮抗的。
模式豆科植物叶片外植体的SE诱导Medicago truncatula(与本研究中使用的芽外植体类似的实验系统)也被用于直接比较反应性和非反应性基因型[72]。蛋白质组学分析显示,在SE诱导处理的第一周内,在一个无反应系中,高水平的蛋白质积累反映了巨大的生理差异,其中一些被确定为应激蛋白。虽然从现有的数据来看,与生物防御激发的关联并不明显,但这使得作者提出,诱导治疗产生的对应激的过度反应可能与缺乏反应有关[73]。这种情况类似于G12外植体的强烈生理反应,并使人联想到上文所讨论的非胚性组织中防御蛋白的高水平积累。
最后,尽管这项研究为生物防御激发和对SE诱导的抗拒之间的联系提供了证据,但它也产生了无法在实验设计中直接解决的假设。首先,只分析了一种无反应基因型,因此其他无反应基因型是否也会发生类似的生理反应仍然没有答案。其次,生物防御激发与SE诱导抑制之间的直接关联的假设仍有待验证。因此,在G6和G12外植体中观察到的生理反应差异可能是与SE反应无关的基因型差异的结果。第三,即使减轻了生物防御诱导,也不清楚G12外植体中是否会诱导SE。因此,在明确了解这些研究结果的广泛适用性和影响之前,还需要进行更多的工作。
结论
本研究的中心结论是,针叶树芽外植体的生理反应,特别是与防御反应的引发有关的生理反应,可能是SE诱导反应的重要决定因素。虽然明确证明生物防御激活对SE诱导是拮抗的还需要更多的证据,但许多一般特征,如与其激发相关的戏剧性代谢和转录重编程,支持了外植体对SE诱导的抵抗作用。此外,它为研究外植体中防御反应的激活和强度的调节机制开辟了新的途径,同时也为开发可用于抑制它们的方法提供了前景,期望这不仅可以产生更有利于SE诱导的生理状态,而且可能对其他类型的组织培养操作更有利。
方法
原始芽的收集和体细胞胚的诱导
2009年5月4日和6日采集了9岁生枝条第二轮和第三轮的芽云杉glauca(白云杉)如前所述由体细胞胚胎产生的树木[9]。其中包括应答型(893-6:G6)和非应答型(893-12:G12),每个基因型共采集了700个无性系树的芽。SE诱导按上文所述进行[9]。简单地说,对芽进行消毒,切除原芽,纵向切成2或4等份,将外植体置于含有9.5 μM 2,4-二氯苯氧乙酸和4.5 μM苄基腺嘌呤的半固体MLV-S培养基上。在培养3、7、15和21天后收集重复外植体样品,液氮冷冻,- 80°C保存。所有剩余的外植体培养16周,在体视显微镜下证实胚团的形成并记录。
RNA制备
进行两轮RNA提取,分别为样品系列1和2。对于用于微阵列分析的样品系列1,采用CTAB-LiCl沉淀方案[74]从这两种基因型的5个生物重复中提取RNA,包括大约80毫克的芽,这些芽在田间收集后(第0天)立即放入液氮中,或在SE诱导处理一周后(第7天)。对于qPCR分析,这些RNA样品的等分在cDNA生成之前进行DNase处理,如反转录部分所述。
对于样品系列2,在诱导的第3天、第15天和第21天采集了3个重复,每个重复由大约80 mg的新鲜质量组成。这些被放置在一个2ml的Sarstedt锥形微管中,其中包含一个5毫米的不锈钢珠,在液氮中冷冻,并在- 80°C下保存。然而,应该注意的是,随后的工作表明,对于某些样品类型,少量(小于50毫克)可以显著提高RNA的质量和数量。将试管转移到适配器组中,该适配器组在- 80°C预冷至少两小时,并在含有几英寸液氮的冷却器中运输,以防止样品解冻。用Qiagen Tissuelyzer II型磨头机在26 Hz下对组织进行两次45 s的破坏。适配器组返回到冷却器,每根管子一次取出一个。每管加入550 μl裂解缓冲液(4 M异硫氰酸胍,0.2 M乙酸钠,pH 5.0, 25 mM EDTA, 2.5% (wt/vol) PVP-40) [75]。试管高速旋转,在56°C下孵育2分钟,在此期间进行一到两个涡流步骤。然后将样品短暂离心以去除细胞碎片,并使用Qiagen RNEasy植物迷你试剂盒(Cat. 10)提取450 μl RNA。# 74904)。使用Qiacube DNA/RNA纯化机器人(Qiagen)进行RNA提取,包括柱上DNA酶处理(Qiagen RNase free DNase, Cat)。# 79254)。然而,随后通过qPCR检测到的基因组DNA数量不同,因此需要进行第二次DNA酶处理(详见反转录部分)。dna酶处理后,使用Nanodrop 1000分光光度计(Thermo Scientific)定量RNA,使用Agilent 2100生物分析仪(Agilent Technologies)评估RNA完整性,其RIN值为8.2-9.8。
微阵列分析
微阵列实验和分析如前所述[12],稍加修改。简单地说,使用Amino Allyl MessageAmpII RNA扩增试剂盒(Life Technologies, Carlsbad, CA, USA)扩增每个总RNA样品1 μg,片段化和定量,并用AlexaFluor 647 (Life Technologies)标记扩增的5 μg RNA。在HS400Pro杂交站(Tecan Group Ltd., Männedorf,瑞士)对寡核苷酸阵列进行预杂交、标记样品与载玻片杂交、载玻片清洗和干燥。切片扫描和特征提取分别在ScanArray Express扫描仪(PerkinElmer, Waltham, MA, USA)和QuantArray v3.0 (PerkinElmer)上完成。每个数组图像分为两部分(上半部分和下半部分)进行分析,并使用内部宏将这两部分融合到Excel中的一个文件中。所有的阵列(20个)都被保留以供进一步分析。实验采用单色设计,4组5组(2个时间点、2个基因型,5个生物重复)。质量控制和数据处理,即背景和缓冲减法,多重测试的分位数归一化和校正(Benjamini-Hotchberg),在R 2.8.1版本中完成[76]。所有微阵列数据已存储在NCBI的基因表达Omnibus中,并可通过GEO系列登录号GSE46977访问。根据调整后的p值选择差异表达基因(p值< 0.05)。选用Flexarray v1.6软件对所选基因进行方差分析和t检验[77]。
候选靶标选择和引物设计
在不考虑靶基因同一性的情况下,根据SE诱导第7天两种基因型之间的表达倍数差异选择候选靶标(表2)2,3.;额外的文件1).第二个考虑因素是在芽采集点(第0天)两种基因型中的相似表达,这导致1个G12和6个G6靶点被排斥。每个候选靶点的探针序列被发射到NCBI上云杉EST库和核苷酸收集数据库,从中构建核苷酸比对以进行引物选择(附加文件)4和5),该方法主要基于将扩增子定位在停止密码子附近,以最小化部分逆转录引起的方差。
引物设计的中心参数是确定足以产生预测T的长度米使用Integrated DNA Technologies在线OligoAnalyzer检测70°C。这是基于通过任意调整Mg来校准程序++参数浓度为50mm,使预测T米cal1f1和cal1r1引物(附加文件5)达到70°C,这种方法在其他寡核苷酸设计程序中被发现是成功的。校准后,通过简单地调整其长度来设计候选引物,直到预测的T米刚刚超过70°C。广泛的自互补引物或与对应引物互补的引物被拒绝,在无模板控制扩增中产生非特异性产物或产生扩增效率<99%的引物对也被拒绝。扩增子大小限制在80-200 bp;然而,预测引物性能的大量努力,如分析引物或扩增子的二级结构,都是不成功的。因此,有必要对某些目标测试多个引物对组合,拒绝那些由LRE分析表明会产生轮廓塌陷或广泛的高原漂移的引物对组合(见[18]查阅有关这些异常的详情)。
反转录
在进行反转录之前,通过扩增20 ng的原始RNA样本来定量基因组DNA污染。这表明许多样品含有少量的dna污染(10-100个基因组),因此所有RNA样品都使用Ambion Turbo无dna dna酶试剂盒(Cat.)进行dna处理。# AM1907),发现它可以将dna污染降低到无法检测的水平。
在含有50 ng/μl RNA、25 ng/μl oligo dT引物(Invitrogen, Cat.)的20 μl反应中进行逆转录。# 18418-012), 5 U/μl上标II (Invitrogen, Cat;# 18064-014),使用制造商提供的缓冲液,在42°C下孵育50分钟,然后加入180 μl 10 mM Tris, pH 8.0,产生最终浓度为5 ng RNA / μl。测试表明,通常采用的RNase抑制剂和DTT的添加没有任何好处。还请注意,我们发现RNase H处理会对一些cDNA靶点产生广泛的复制谱散射,这大大降低了定量准确性。如前所述,使用相同RNA样本进行三次重复逆转录酶反应,对三个参考基因靶点产生约±12%的平均方差[18],表明该方法具有高度重复性,与本研究中观察到的内参基因表达的小方差一致(见下文)。
LRE qPCR
LRE qPCR是如何开发的详细描述,它的性能能力,以及自动化LRE分析的独立于平台的Java桌面程序的描述,先前已发表[16- - - - - -18,78]。简而言之,绝对定量是通过将每个扩增图谱中心区域的3-6个荧光读数转换成以荧光单位表示的目标量(F0).这些被平均并转换成DNA质量(M)0)使用由已知数量的lambda基因组DNA扩增产生的光学校准因子(OCF =每ng dsDNA的荧光单位),这种方法类似于使用荧光染料对核酸进行定量。然后将其转化为目标分子数(N)0),基于放大子尺寸(A年代) [17]:
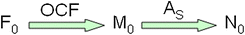
PCR扩增采用Applied Biosystems 7500 Fast qPCR系统(正常提速),定量酶制剂(Qiagen, Cat);# 204145),反应体积为10 μl,含有500 nM引物,相当于5 ng RNA的逆转录酶反应,使用96孔BrightWhite板(Primerdesign, BW-Fast),密封MicroAmp膜(Applied Biosystems, Cat # 4311971)。循环过程包括在95°C下激活15分钟,然后在95°C下循环10秒,在65°C下循环120秒。扩增子T米在每次运行结束时,通过熔体曲线分析(65至90°C)来确定每个扩增反应。原始荧光读数被导入到LRE Analyzer (Version 0.8.7)中,这是一个开源Java程序,可在此下载[79,80]。该程序包含一个广泛的帮助数据集,描述了该程序的功能,以及设置和测试qPCR分析性能的各种指南。
熔体曲线分析显示,所有引物对都产生一个突出的扩增峰,扩增效率在102 - 107%之间。如LRE分析仪帮助中所述,quantiect产生大于100%的扩增效率的倾向,即产生适度低估目标数量的系统性偏差,通过将扩增效率固定为100%来补偿。LRE Analyzer数据库在附加文件中提供6,以及本研究中使用的放大子和光学校准数据库。个别定量测定的摘要在附加文件中提供7。
内参基因分析
9个内参基因的表达被用来确定生物变异水平,除了作为内部质量控制来评估与样品制备和LRE qPCR分析相关的技术差异。从微阵列分析中提取了两个内参基因(假设蛋白(HP)和过氧化物酶体靶向信号受体(PTSR)) [81],其余7个与拟南芥内参基因同源的针叶树基因也通过微阵列分析鉴定出来[82]。引物序列连同UniGene加入编号在附加文件中提供5。
评估表达稳定性基于变异系数,类似于开发genevinvestigator’s RefGenes工具的方法,其中使用微阵列分析产生的信号强度的标准差来评估转录组全表达稳定性[83]。数字6A提供了基于样品系列1(第0天和第7天)中EF1α表达的该方法的示例。组内方差是生物变异和与RNA制备、cDNA生成和LRE qPCR分析相关的技术衍生方差的组合,而组间方差主要反映生物变异。将分析扩展到9个内参基因,得到了相似的组内方差(图2)6B),组间差异更大(图2)6C)。

样品系列1内参考基因表达分析:第0天和第7天。(一)基于EF1α表达的变异系数(±CV)确定组内和组间方差的代表性示例。(B)9个内参基因的组内方差。(C)基于平均转录本数量的组间差异见(一)。数量(一)和(C)表达为每10 ng RNA的转录本。±CV =(标准差/平均值)× 100%。
对样本系列2重复分析(第3天、第15天和第21天)产生了类似的组内变化(图2)7A)。除CDC2和UBC1外,组间差异也相似(图2)7B),远低于样本系列1的观测值(图2)6C)。另一个值得注意的结果是,尽管具有高水平的表达稳定性,但样品系列1中几乎所有内参基因的平均绝对数量都低于样品系列2(图2)7C)。虽然没有研究这些差异的来源,但可能与样品系列1中用于制备RNA的LiCl沉淀步骤有关。无论如何,基于这样一个前提,即这种异常相对于在候选基因表达中观察到的巨大变化是适度的,并且它只会影响第0天和第7天的样本,对于本研究的目的,这种定量偏差被认为是微不足道的。总体而言,这些大型数据集表明,除了说明LRE qPCR可以实现的定量精度外,所有10个样本组的生物学变异性水平非常低。

样品系列2内参考基因表达分析:第3、5、21天。(一)组内变异(±CV)。(B)基于平均转录本数量的组间差异如图所示6一个。(C)从样本系列1和2得出的平均绝对数量的差异。所有数量都以每10 ng RNA的转录物表示。±CV =(标准差/平均值)× 100%。
参考文献
植物组织培养,自Gottlieb Haberlandt以来的100年。观赏植物的微繁。编辑:Laimer M, r
Nehra NS, Becwar MR, Rottmann WH, Pearson L, Chowdhury K, Chang S, Dayton Wilde H, Kodrzycki RJ, Zhang C, Gause KC, Parks DW, Hinchee M:体外细胞发育生物学。森林生物技术:创新方法,新机会第41版。2005年,701 - 717。
李建军,李建军,李建军,李建军,等。松木体胚发育的研究进展。植物生理学报,2007,29(1):444 - 444。
彭建军,陈建军,陈建军,等。克隆繁殖对松柏类植物抗性的影响。植物细胞与组织器官学报,2010,31(2):591 - 594。10.1007 / s11240 - 009 - 9647 - 2。
杨建军,李建军,李建军,李建军,李建军,李建军,李建军,李建军,李建军,李建军,李建军,李建军,等。黑松种子体细胞胚发生的初步研究进展。植物物理学报,2010,30:1469-1478。10.1093 / treephys / tpq081。
李建军,李建军,李建军,等。植物干细胞的多能性研究进展[j]。植物科学进展,2007,12:245-252。10.1016 / j.tplants.2007.04.002。
张宏,Ogas J:种子基因发育调控的表观遗传学研究。植物学报,2009,26(2):610-627。10.1093 / mp / ssp027。
杨建军,杨建军,李建军,等。植物干细胞在逆境胁迫下的发育与调控。植物科学,2011,22(2):1-10。
Klimaszewska K, Overton C, Stewart D, Rutledge RG: 10年白云杉体细胞胚的形成和植株再生及其组织培养过程中11个基因的表达谱。植物学报,2011,33(3):635-647。10.1007 / s00425 - 010 - 1325 - 4。
李建军,李建军,李建军,李建军,李建军,李建军,李建军,李建军。植物生理学报,2011,37(4):557 - 557。10.1104 / pp.111.179663。
MacKay JJ, Dean JFD, Plomion C, Peterson DG, Cánovas FM, Pavy N, Ingvarsson PK, Savolainen O, Guevara MA, Fluch S, Vinceti B, Abarca D, Díaz-Sala C, Cervera MT:解码针叶树千兆基因组。植物生理学报,2012,31(6):555-569。10.1007 / s11103 - 012 - 9961 - 7。
Raherison ES, Rigault P, Caron S, Poulin PL, Boyle B, Verta JP, gigu
杨建军,陈建军,陈建军,陈建军,等。挪威云杉花蕾萌发时间与脱氢酶基因表达的关系。植物学报,2008,28(2):459-472。10.1007 / s00425 - 008 - 0750 - 0。
张建军,张建军,张建军,等。信号肽的分离与分析。科学通报,2011,(8):785-786。10.1038 / nmeth.1701。
Silverstein KT, Moskal W, Wu HC, Underwood B, Graham M, Town CD, VandenBosch K:植物中类似抗菌肽的富含半胱氨酸的小肽被低估了。植物科学学报,2007,31(1):369 - 369。10.1111 / j.1365 - 313 x.2007.03136.x。
李建军,李建军,李建军,等。聚合酶链反应的s型动力学模型及其在高容量实时定量PCR中的应用。中国生物医学工程学报,2008,28(8):457 - 457。
Rutledge RG, Stewart D:评估基于lre的绝对定量实时PCR检测的性能。PLoS ONE。2010, 5: e9731-10.1371/journal.pone.0009731。
Rutledge RG:基于lre的实时qPCR Java程序,可实现大规模绝对定量。PLoS ONE。2011, 6: e17636-10.1371/journal.pone.0017636。
范德松,王晓明,王晓明,等。实时荧光定量PCR技术的研究进展。参考基因验证软件改进的标准化第4版。编辑:洛根J,爱德华兹K,桑德斯NB。2009,英国诺福克:卡斯特学术出版社,47-64页。
Schmittgen TD, Livak KJ:比较CT法分析实时PCR数据。Nat协议,2008,32(3):1101-1108。10.1038 / nprot.2008.73。
Huggett JF, Dheda K, Bustin SA:实时RT-PCR归一化;策略和考虑。中国生物医学工程学报,2005,6(6):779 - 784。10.1038 / sj.gene.6364190。
古铁雷,李建军,李建军,李建军,李建军,李建军,李建军,李建军,李建军,等。植物内参基因的系统验证:逆转录-聚合酶链反应(RT-PCR)分析中存在的严重缺陷。植物营养学报,2009,29(6):693 - 698。10.1111 / j.1467-7652.2008.00346.x。
植物抗病性中的氧化爆发。植物生理学报,1997,28(4):551 - 557。10.1146 / annurev.arplant.48.1.251。
Alvarez M, penell RI, Meijer P-j, Ishikawa A, Dixon RA, Lamb C:活性氧中间体介导植物免疫系统信号网络的建立。细胞,1998,32(2):773-784。10.1016 / s0092 - 8674(00) 81405 - 1。
马瑞诺D,杜楠C, Puppo A,保利N:植物NADPH氧化酶的爆发。植物科学,2012,17:9-15。10.1016 / j.tplants.2011.10.001。
张永华,常红生,古普瑞,王晓,朱婷,栾生:拟南芥损伤、病原体、非生物胁迫和激素应答之间的转录分析。植物生理学报,2002,29(2):661-677。10.1104 / pp.002857。
张建军,张建军,张建军,等。生物信息学研究进展。植物生理学报,2006,41(1):373-378。10.1104 / pp.106.079467。
李建军,李建军,李建军,等。茉莉素诱导基因的研究进展。植物科学进展,2009,14:87-91。10.1016 / j.tplants.2008.11.005。
Chaouch S, Queval G, Noctor G: AtRbohF是拟南芥防御相关代谢的重要调节因子,是细胞内氧化应激与发病反应相互作用的关键因子。植物科学学报,2012,32(1):613-627。10.1111 / j.1365 - 313 x.2011.04816.x。
Robert-Seilaniantz A, Grant MR, Jones JDG:植物病害和防御中的激素串扰:不仅仅是茉莉酸-水杨酸拮抗剂。植物保护学报,2011,49(4):317-343。10.1146 / annurev -发朵- 073009 - 114447。
王志强,王志强,王志强,等。植物激素相声调控的研究进展。植物科学进展,2012,17:22-31。10.1016 / j.tplants.2011.10.006。
张建军,张建军,张建军,等。茉莉酸盐和水杨酸盐信号串扰的研究进展。植物科学,2012,17:260-270。10.1016 / j.tplants.2012.02.010。
王晓东,王晓东,王晓东,等。活性氧:代谢、氧化应激和信号转导。植物生态学报,2004,25(5):373- 379。10.1146 / annurev.arplant.55.031903.141701。
Passardi F, Cosio C, Penel C, Dunand C:过氧化物酶的功能比瑞士军刀还多。植物学报,2005,24(4):559 - 564。10.1007 / s00299 - 005 - 0972 - 6。
Almagro L, Gómez Ros LV, Belchi-Navarro S, Bru R, Ros Barceló A, Pedreño M: III类过氧化物酶在植物防御反应中的作用。[J] .中国生物医学工程学报,2009,35(6):377- 391。10.1093 / jxb / ern277。
Cosio C, Dunand C:个体III类过氧化物酶基因的特定功能。中国生物医学工程学报,2009,35(6):991 - 998。10.1093 / jxb / ern318。
Doehlemann G, Linde KVD, Aßmann D, Schwammbach D, Mohanty A, Jackson D, Kahmann R:麦氏黑穗病菌(Ustilago maydis)分泌的效应蛋白Pep1是成功入侵植物细胞所必需的。中华医学杂志,2009,5:e1000290-10.1371/journal.ppat.1000290。
Hemetsberger C, Herrberger C, Zechmann B, Hillmer M, Doehlemann G:黑穗病菌效应物Pep1通过抑制宿主过氧化物酶活性来抑制植物免疫。中华医学杂志,2012,8:1002684-10.1371/journal.ppat.1002684。
kkjaersg
黄文杰,黄文杰,黄文杰。表皮缺陷导致对一种主要植物病原体的完全免疫。植物学报,2007,49(4):972-980。10.1111 / j.1365 - 313 x.2006.03017.x。
李建军,李建军,李建军,等。植物胁迫生物学中细胞氧化还原状态的研究进展。植物生理学报,2010,48(4):292-300。10.1016 / j.plaphy.2009.12.007。
王晓明,王晓明,王晓明,王晓明,王晓明,王晓明,王晓明,王晓明,王晓明,王晓明,王晓明,王晓明,王晓明,等。植物科学进展,2011,16:300-309。10.1016 / j.tplants.2011.03.007。
Foyer CH, Noctor G:抗坏血酸和谷胱甘肽:氧化还原中枢的心脏。植物生理学报,2011,32(5):518 - 518。10.1104 / pp.110.167569。
李建军,李建军,李建军,李建军。植物脱水酶基因的克隆及其在逆境胁迫中的应用。植物信号学报,2011,6(6):1503-1509。10.4161 / psb.6.10.17088。
李建军,李建军,李建军,等。细胞外转化酶的研究进展。[J] .中国生物医学工程学报,2013,35(4):513-524。10.1093 / jxb / erg050。
Roitsch T, González MC:植物转化酶的功能和调控:甜味感觉。植物科学进展,2004,9:606-613。10.1016 / j.tplants.2004.10.009。
植物转化酶:结构、功能和调控。中国生物医学工程学报,2005,31(4):369 - 369。
李春华,李春华,李春华,等。植物生理学与植物病理学:植物初级代谢与植物-病原体相互作用。[J] .实验学报,2007,58(5):419 - 426。10.1093 / jxb / erm298。
博尔顿博士:初级代谢和植物防御——火灾的燃料。植物学报,2009,22(2):487-497。10.1094 / mpmi - 22 - 5 - 0487。
王晓明,王晓明,王晓明,等。RNA干扰对烟草源叶细胞壁转化酶表达的影响。植物生理学报,2009,32(1):387 - 398。10.1104 / pp.108.121418。
范龙,李建军,李建军,等。植物抗侵染诱导防御相关蛋白的研究进展。植物酚类植物学报,2006,44(4):135-162。10.1146 / annurev.phyto.44.070505.143425。
Chou HM, Bundock N, Rolfe SA, Scholes JD:拟南芥叶片感染念珠菌(白色水疱锈病)导致宿主代谢重编程。植物病理学杂志,2000,1:99-113。10.1046 / j.1364-3703.2000.00013.x。
Fotopoulos V, Gilbert MJ, Pittman JK, Marvier AC, Buchanan AJ, Sauer N, Hall JL, Williams LE:单糖转运基因AtSTP4和细胞壁转化酶Atbetafruct1在拟南芥感染真菌生物营养菌Erysiphe cichoracearum后的诱导作用。植物生理学报,2003,32(2):821-829。10.1104 / pp.103.021428。
Bonfig KB, Schreiber U, Gabler A, Roitsch T, Berger S:侵染对拟南芥叶片光合作用和库代谢的影响。植物学报,2006,25(2):1-12。10.1007 / s00425 - 006 - 0303 - 3。
李建军,陈建军,陈建军,陈建军,陈建军。拟南芥转录组对根癌农杆菌转录组的响应。植物学报,2006,19(2):665-681。10.1094 / mpmi - 19 - 0665。
Quilliam RS, Swarbrick PJ, Scholes JD, Rolfe SA:拟南芥损伤叶片光合作用成像。[J] .中国生物医学工程学报,2006,27(5):555 - 569。
罗林斯,李建平,张建平,等。多肽酶抑制剂的研究进展。中国生物医学工程学报,2004,31(2):559 - 564。10.1042 / BJ20031825。
张建军,张建军,张建军,等。蛋白质水解酶的研究进展。中国生物医学工程学报,2012,35(4):343- 356。10.1093 / nar / gkr987。
Silverstein KAT, Graham MA, Paape TD, VandenBosch KA:拟南芥中300多个防御样基因的基因组组织。植物生理学报,2009,28(3):693 - 693。10.1104 / pp.105.060079。
李建军,李建军,李建军,等。植物丝氨酸蛋白酶抑制剂的研究进展。植物信号学报,2011,6(6):109 -1011。10.4161 / psb.6.7.15504。
Morán-Diez E, Rubio B, Domínguez S, Hermosa R, Monte E, Nicolás C:拟南芥与生防真菌哈兹木霉培养24 h后的转录组反应。植物生理学报,2012,32(1):557 - 557。10.1016 / j.jplph.2011.12.016。
Denoux C, Galletti R, Mammarella N, Gopalan S, Werck D, De Lorenzo G, Ferrari S, Ausubel FM, Dewdney J:拟南芥幼苗OGs和Flg22诱导子的防御反应通路激活。植物学报,2008,31(1):423-445。10.1093 / mp / ssn019。
Prins A, Mukubi JM, Pellny TK, Verrier PJ, Beyene G, Lopes MS, Emami K, Treumann A, llarge - trouverie C, Noctor G, Kunert KJ, Kerchev P, Foyer CH:叶片等级对玉米高CO2适应的影响。植物学报,2011,34(4):314-331。10.1111 / j.1365-3040.2010.02245.x。
李春华,王晓明,王晓明,等。转基因拟南芥植物对氧化胁迫的抗性研究。植物生理学报,2008,28(4):369 - 369。10.1104 / pp.108.124875。
范胡腾M, Pelser M, Van Loon LC, Pieterse M, Ton J:拟南芥启动防御的成本和效益。美国国家科学促进会。[j] .中国科学:地球科学。10.1073 / pnas.0510213103。
王晓明,王晓明,王晓明,等。拟南芥对水杨酸的抗性与生物量的关系。植物学报,2010,32(2):444 - 444。10.1111 / j.1467-7652.2009.00468.x。
李建军,李建军,李建军,李建军,李建军。拟南芥细胞中茉莉酸甲酯介导的转录重编程和细胞周期的研究进展。科学通报,2008,35(5):990 - 998。10.1073 / pnas.0711203105。
马绍宁,李春华,李春华,李春华,李春华。葡萄体细胞胚胎发生的蛋白质组学分析。植物学报,2008,27(2):347-356。10.1007 / s00299 - 007 - 0438 - 0。
张军,马宏,陈生,季敏,Perl A, Kovacs L,陈生:葡萄胚性和非胚性愈伤组织中胁迫响应蛋白的差异表达。赤霞珠-蛋白质组学方法。植物科学,2009,17(1):103-113。10.1016 / j.plantsci.2009.04.003。
王晓明,王晓明,王晓明,等。藏红花胚性愈伤组织与非胚性愈伤组织相关蛋白的克隆。中国生物医学工程学报,2012,32(1):1 - 4。
Correia S, Vinhas R, Manadas B, loureno AS, Veríssimo P, Canhoto JM:生长素诱导的茄科树Cyphomandra betacea (Tamarillo)胚性和非胚性组织的比较蛋白质组学分析。中国生物医学工程学报,2012,31(2):444 - 444。10.1021 / pr200856w。
Rose RJ, Nolan KE:体外细胞发育生物学。特邀综述:体细胞胚胎发生的遗传调控,特别是拟南芥和苜蓿42版。2006年,473 - 481。
Almeida AM, Parreira JR, Santos R, Duque AS, Francisco R, tom
张建军,张建军,张建军,张建军,张建军,张建军,张建军,张建军,张建军,张建军,张建军,张建军,张建军,张建军,张建军,张建军,张建军。中国生物医学工程学报,2008,31(2):766-786。10.1111 / j.1469-8137.2008.02615.x。
李建军,李建军,李建军,等。基于逆转录聚合酶链反应的木本植物RNA提取技术及其应用。植物学报,1997,31(1):229 -226。10.1094 / PDIS.1997.81.2.222。
R核心团队:用于统计计算的语言和环境。http://www.R-project.org,
FlexArray:用于基因表达微阵列的统计数据分析软件。麦吉尔大学和魁北克基因组创新中心,http://www.gqinnovationcenter.com/services/bioinformatics/flexarray/index.aspx?l=e,
Rutledge RG, Stewart D:用于确定扩增效率的方法的关键评估驳斥了实时PCR的指数特征。中国生物医学工程学报,2008,29(9):996 - 996。
LRE qPCR网站。http://sites.google.com/site/lreqpcr/,
LRE qPCR分析仪。http://code.google.com/p/lreqpcr/,
Friedmann M, Ralph SG, Aeschliman D, Zhuang J, Ritland K, Ellis BE, Bohlmann J, Douglas CJ:中国云杉(Picea sitchensis)顶芽发育过渡基因的微阵列表达谱分析。[J] .中国生物医学工程学报,2007,35(5):593- 594。
赵建军,李建军,李建军,等。拟南芥转录调控基因的全基因组鉴定和内参基因的检测。植物生理学报,2005,39(5):517 - 517。10.1104 / pp.105.063743。
Hruz M, Wyss M, Docquier M, Pfaffl MW, Masanetz S, Borghi L, Verbrugghe P, Kalaydjieva L, Bleuler S, Laule O, Descombes P, Gruissem W, Zimmermann P: RefGenes: RT-qPCR数据归一化的可靠内参基因。中国生物医学工程学报,2011,32(2):559 - 564。
致谢
作者感谢A. ssamguin对手稿的严格审查和I. Lamarre的编辑协助。这项工作的资金由加拿大自然资源部(RGR, KK)、加拿大基因组部、魁北克基因组部、加拿大基因组部为Arborea和SMarTForests项目(JM)以及自然科学与工程委员会(JM)的发现资助计划提供。
作者信息
从属关系
相应的作者
额外的信息
相互竞争的利益
作者没有非金融、金融或专利相关的竞争利益。
作者的贡献
KK、SC、BB和JM设计实验方法,KK和CO采集芽并进行SE诱导,SC和BB进行芯片分析,DS和RGR进行qPCR基因表达分析,RGR撰写了KK、SC、BB和JM的稿件。所有作者都阅读并批准了最终的手稿。
权利和权限
本文由BioMed Central Ltd.授权发表。这是一篇基于知识共享署名许可(http://creativecommons.org/licenses/by/2.0),允许在任何媒体上不受限制地使用、分发和复制,前提是正确引用原创作品。
关于本文
引用本文
拉特利奇,r.g.,斯图尔特,D.,卡伦,S.。et al。白云杉成树芽原基诱导体细胞胚胎发生的生物防御激活与抵抗之间的潜在联系(云杉glauca).BMC Plant Biol13日,116(2013)。https://doi.org/10.1186/1471-2229-13-116
收到了:
接受:
发表:
DOI:https://doi.org/10.1186/1471-2229-13-116
关键字
- 针叶树
- 基因表达谱
- 微阵列分析
- 绝对qPCR
- LRE qPCR
- 无性繁殖