
摘要
背景
microrna (mirna)和短干扰rna (sirna)在生物应激反应(如病毒感染)中的作用已经在植物中得到了证实。番茄黄曲叶病毒(TYLCCNV)是一种具有系统性侵染能力的单组分begomavirusSolanaceae.植物,并诱导叶片卷曲,燃烧和致病症状,当与β外晶体(Tylccnb)共同接种。释放的基因组序列尼古利亚娜·宾夕法尼亚州为鉴定对begomovirus相关的betat卫星应答的mirna和sirna提供了机会N. Benthamiana..
结果
在3个小RNA库中鉴定了mirnaN. Benthamiana.单独用Tylccnv(Y10A)系统感染的植物,共同感染Y10a及其β脱肽Tylcnb(Y10β)或含有突变的Tylccnb突变体(Y10Mβ)βC1,唯一的β卫星编码蛋白。从160个家庭的38个家庭和197个新的MiRNA共有196家保守的MiRNA被确定。Northern印迹分析证实,特异性特异性miRNA的表达远低于保守的miRNA。发现几个保守的和新的miRNA对Y10a和Y10β的共感染,但不对Y10a和Y10Mβ的共同感染,这表明这些miRNA可能在Y10β和Y10β之间相互作用起作用起作用。N. Benthamiana..此外,我们还发现了可以触发分阶段次级sirna (phasiRNAs)产生的mirna。
结论
鉴别鉴别差异表达概况的miRNAN. Benthamiana.同时感染Y10A和Y10β以及同时感染Y10A和Y10mβ表明这些mirna是β卫星应答的。我们的结果也提示了mirna介导的phasirna的产生在begomovirus和N. Benthamiana..
背景
由Micrornas(miRNA)和短干扰RNA(siRNA)组成的内源性小RNA是器官发育,应力反应和植物基因组稳定性的重要调节因子[1,2].MiRNA和SiRNA在生物和非生物应激反应中的角色,例如病毒感染,许多研究已经证明了植物中的[2].在病毒感染的植物中mirna的下调或上调,如miR156、miR158、miR159、miR160、miR164、miR168、miR171、miR172、miR319、miR398和miR1885,可能在病毒与植物的相互作用中发挥作用[3.- - - - - -10.].植物内源性sirna介导的基因沉默与细菌诱导的植物生物胁迫有关nat-siRNAATGB2调节R-基因介导的植物免疫[11.].水稻条纹病毒感染提高了序列MiRNA从特定前体的积累[12.].
植物mirna可以触发非编码(如:TAS.)或蛋白质编码基因(例如NBS-LRR基因)。反式作用sirna (tasiRNAs)是一种独特的sirna,它是由TAS.与miRNA切割位点相关的21核苷酸(nt)相转录本。他们的行为在反式调节在后术语水平的基因表达。通过前体的单一或双端的miRNA的相互作用来触发TASIRNA的生物发生TAS.转录物[13.].至少有八个TAS.轨迹属于四个TAS.家庭(TAS1.- - - - - -TAS4.)拟南芥四TAS3.稻米已识别出稻米[13.- - - - - -15.].最近的研究表明,植物基因组富含序列的siRNA(PHASiRNA) - 发布基因座,或pha基因[16.- - - - - -19.,并且可能在蛋白质编码基因中含有数百个这样的位点[15.,20.- - - - - -23].例如,在水稻花序发育过程中发现了一批由miR2118和miR2275分别触发的21 nt和24 nt phasirna [18.].phasirna的生物发生及其在转录后调节中的作用在最近的综述中得到了很好的讨论[24].
双生病毒是一组单链DNA病毒,在世界范围内造成毁灭性的经济损失。四个属(mastrevirus.,Begomovirus.,Curtovirus和Topocuvirus)根据基因组组织,主机范围和传输向量进行分类Begomovirus.数量最多,地理分布最广的[25].前豚病毒有单氨酸或二分离基因组。在过去的十年中,几项研究表明,许多单牛石病毒血管病毒已经获得了βatellite,以前称为DNAβ,和βC1,唯一的β丁钛合体编码的蛋白质在症状诱导中起重要作用,抑制转录和后术基因沉默,并且还涉及抑制所选的茉莉酸反应[25- - - - - -29].番茄黄叶卷曲中国病毒(Tylccnv)是与βatellite(Tylccnb)相关的单牛石豚霉狼。单独的tylcnv产生了无症状的感染尼古利亚娜·宾夕法尼亚州和番茄植物。TYLCCNV与这些寄主植物共感染TYLCCNV时,TYLCCNB是诱导叶曲症状的必要条件。TYLCCNB突变体无法表达βC1蛋白质在接种TylCCNV接种时,蛋白质废除了诱导植物中典型症状的能力[27].在我们之前的研究中,我们通过对系统性感染的小rna进行深度测序,识别出TYLCCNV-和tylccnb来源的sirnaN. Benthamiana.发现病毒转录物可能作为RNA依赖的RNA聚合酶(RDR)底物而产生二级siRNA [30.].然而,TYLCCNB调控的植物miRNA谱仍然知之甚少。最近发布的基因组序列草案N. Benthamiana.[31[响应病毒感染,提供从宿主植物中鉴定miRNA和siRNA的机会,并研究宿主miRNA和siRNA的表达变化。在这项研究中,我们确定了N. Benthamiana.MIRNAS基于GRAF GRAF组和我们先前生成的小型RNA数据集使用N. Benthamiana.感染的植物感染了Begomovirus,并发现许多麦芽瘤,包括少量推定的阶段触发器,对TylCCNV和TylCCNB的共感染而不是Tylccnv和突变的TylccnbN. Benthamiana..
结果
miRNAs的鉴定N. Benthamiana.
的N. Benthamiana.单独感染TYLCCNV (Y10A) (P1)或联合感染TYLCCNV及其betassatellite TYLCCNB (Y10β;P2)或TYLCCNB突变体(Y10mβ;P3),用于RNA提取和小RNA测序[30.].在1220万小RNA中阅读或730万独特的阅读N. Benthamiana.叶子,总计720万读或5.6万张独特的读数可以映射到N. Benthamiana.基因组(附加文件1:表S1)。采用上述方法,利用BLAST对公众miRNA数据库(miRBase, Release 19)进行搜索,我们共鉴定了来自38个家族的196个保守miRNAN. Benthamiana.基因组(附加文件2:表S2)。同时,还确定了来自160个家庭的197个新的MiRNA(附加档案3.:表S3)。进一步的搜索(Blastn与<1e-3)这些新的成熟miRNA序列来自小RNA种群n .烟草最近由我们产生[32]表明了42个小说N. Benthamiana.miRNAs是保守的n .烟草.目标预测指示147(91.9%)小型miRNA具有至少一个潜在目标(附加文件3.:表S3)。Northern blot检测到两种新的mirna Nbe-miR70和Nbe-miR145以及保守的miR159的表达,但这两种mirna的表达水平远低于miR159(图)1),与众所周知的低表达非保守miRNAs现象一致。

小说尼古利亚娜·宾夕法尼亚州TYLCCNV/TYLCCNB感染应答的mirna。(一种)和(B)两种小说的前体N. Benthamiana.mirna Nbe-miR70和Nbe-miR145。(C)NBE-MIR70和NBE-MIR145表达的Northern印迹分析N. Benthamiana.TYLCCNV单独感染的植株(Y10A或P1),与TYLCCNB (Y10β或P2)或突变的TYLCCNB (Y10mβ或P3)一起感染。保守miR159作为miRNA对照。
识别推定的临时触发器
根据人类基因组草图N. Benthamiana.以及Howell等人描述的算法。[17.](详情见方法),我们发现超过数千个位点能够生成21 nt和24 nt相rna(相评分> 1.4)。在这些phasiRNA位点中,至少157个21 nt和296个24 nt的phasiRNA位点(在有阶段性小RNA reads的区域及其两侧200 nt区域内)可以预测到miRNA结合位点,表明这些miRNA是推测的phasiRNA触发位点N. Benthamiana.(附加文件4:表S4)。对于这些潜在的阶段触发或阶段寄存器,其推定的裂解位点(在预测的miRNA靶位点中的位置10和11之间)均位于相位位置。共有24个家庭共有62个保守的miRNA,可以结合起来pha找到基因座,例如mir156和mir482(表1).超过300phaLoci似乎是由N. Benthamiana.要么尼古利亚娜- 特价miRNA,例如NBE-MIR2(附加文件4:表S4)。一个例子如图所示2,其中nbe-mir113,一部小说N. Benthamiana.预计miRNA,以触发21-nt相位的小rnas的产生N. Benthamiana.基因。除了非编码的转录本,如TAS.许多蛋白编码基因也被鉴定为mirna触发基因pha位点在N. Benthamiana.(附加文件4:表S4),包括由保守的miRNA定位的一些基因座,例如MiR156和MiR482(表1)已显示在最近的几项研究中从蛋白质编码基因中显示出在蛋白质编码基因中产生phasirnas [15.,20.- - - - - -23,33,34].当施加极其严格的相位评分(> 20)时,仍然可以识别出分别产生21和13个基因座和24-NT Phasirnas。

nbe-mir113触发了Phasirna生产NBS-LRR基因(Niben044Scf0002438)N. Benthamiana..(一种)miRNA与其目标之间的对齐。两个点表示匹配。(B)小型RNA丰富和跨越Niben0444444438的分阶段分量分布。
MiRNA的表达谱响应Tylcnv / Tylcnb的共感染
以前的报告显示,在病毒感染后,一些miRNA差异表达[8,10.].In this study, our focus was on the miRNAs responsive to betasatellite (TYLCCNB) associated with TYLCCNV, i.e. those with a similar expression level in plants infected with TYLCCNV (Y10A) and co-infected with TYLCCNV and a mutated TYLCCNB (i.e. Y10A + Y10mβ) but with an up- or down-regulated expression level in plants co-infected with TYLCCNV and a functional TYLCCNB (i.e. Y10A + Y10β). With the criteria of 1.7-fold changes in Y10A + Y10β versus both Y10A and Y10A + Y10mβ and RPM (reads per million) ≥ 10 in at least one of the three samples, we found that among the conserved miRNAs, miR160, miR164, miR397 and miR398 were up-regulated whereas miR169, miR391 and miR827 were down-regulated in responsive to TYLCCNB, and that among the novel miRNAs, Nbe-miR8, Nbe-miR70, Nbe-miR107, Nbe-miR124 and Nbe-miR134 were up-regulated whereas Nbe-miR2, Nbe-miR3 and Nbe-miR142 were down-regulated in responsive to TYLCCNB (Table1;附加文件5:表S5)。northern blot证实Nbe-miR70上调(图)1).此外,在植物中观察到的植物中观察到的MiR4376的下调和功能性TylCCNB,即Y10A +Y10β,北印迹结果也证实;因此,其靶基因的上调ACA10(自动禁止加利福尼亚州2+腺苷三磷酸酶10),采用RT-qPCR检测(图3.;附加文件5:表S5)。

miR4376及其靶标的表达(ACA10)三种不同cDNA文库的mRNAN. Benthamiana..(一种)miR4376与其靶标对齐ACA10.(B)miR4376在N. Benthamiana.TYLCCNV单独感染的植株(Y10A或P1),与TYLCCNB (Y10β或P2)或突变的TYLCCNB (Y10mβ或P3)一起感染。(C)的表达ACA10在通过实时QPCR测量的各种处理中。误差栏代表用三个生物重复确定的标准偏差。
大多数miRNA都有类似的表达水平N. Benthamiana.单独用TylCCNV系统感染的植物(P1)并与Tylccnv和突变蛋白脂肪体Tyccnb(P3)共感染(附加档案5:表S5),确认无效的βC1在突变的TYLCCNB中。这一结果也表明,上述对β -卫星反应的mirna很可能确实在begomovirus和begomovirus之间的相互作用中发挥作用N. Benthamiana..有趣的是,一些差异表达的mirna,如miR391、miR397和miR398,被认为是phasiRNA产生的触发器(见表)1).
讨论
近期研究证明了MIRNA在不同的生物反应(病毒,细菌和真菌)中的参与[2].我们的研究证实,宿主mirna(如miR391、miR397和miR398)的表达受到β卫星的影响。至少有两种不同的miRNAs:一种被RNA沉默的病毒抑制子(vrs)抑制,另一种通过调节植物固有免疫受体或激素信号通路参与抗病原体(病毒)。病毒编码蛋白质干扰宿主RNAi通路,从而扭曲正常的细胞活动[8].它们可以直接与宿主miRNA结合,从而使RNA诱导的沉默复合物(RISC)的非功能性。miRNA是植物中不同生物过程中的关键调节分子,例如MiR164,MiR159 / 319用于叶片发展[1].显然,典型的叶卷曲症状N. Benthamiana.TYLCCNV和它的betassatellite联合感染诱导的TYLCCNB可能是由那些对betassatellite响应的差异积累的mirna的调节引起的(表1)1).差别下调的miR4376N. Benthamiana.预测TYLCCNV和TYLCCNB共同感染的植物,能靶向aACA基因,也可以参与叶发育,因为最近的一项研究表明MiR4376目标ACA10基因在番茄生殖增长中发挥着关键作用[22].此外,在TYLCCNV和一个突变TYLCCNB共同感染的植物中上调的mirna,如miR160,其上调水平远小于TYLCCNV和一个功能性TYLCCNB共同感染的植物,也可能在响应betassatellite中发挥作用。
最近的研究表明mirna介导的phasiRNA通路在多种生物过程中发挥作用,包括病原体抗性[15.,20.- - - - - -23,34].例如,mir482 /2118介导的抗病分裂NBS-LRR基因不仅在非种族特异性疾病抗性中发挥重要作用,而且还触发了阶段性小rna的产生,这些小rna能够调节其靶蛋白的表达反式[15.,21].在病原体感染后,miR482的表达水平下调,因此其目标,NBS-LRR基因上调[21,34].miR482/2118属于一种特定类型的miRNA,由包含在miRNA/miRNA*双工中的不对称凸体的pre-miRNA产生,并已被证明是通过RDR6/DCL4途径从其靶转录本产生阶段性21-nt次级小rna的触发器[34].因此,预期患有靶疾病抗性基因的MiR482 / 2118介导的靶抗病基因的切割,导致其靶MRNA的衰减和从靶基因中产生分阶段的二次小RNA,例如番茄,棉花和棉花和Medicago truncatula[15.,21,34].此外,从miR482产生的次要小rna中至少有一个靶向NBS-LRR已显示基因对编码另一种无国防相关蛋白的mRNA进行靶向mRNA [21,表明由蛋白质编码基因产生的次级小rna的行为类似TAS.位点在拟南芥[34].在本研究中,miR482表达水平上调N. Benthamiana.与单独感染TylCCNV感染的植物相比,与Tylccnv和Tylccnb共同感染的植物与先前的观察相比。同时,在与TylCCNV和突变的TylCCNB共同感染的植物中也观察到miR482的这种上调(附加文件5:表S5)。因此,miR482对betassatellite的响应功能还有待进一步研究。除了miR482,其他几个β卫星响应的mirna,如miR397和miR398,是phasirna的潜在触发器(附加文件4:表S4),表明这些miRNA和这些miRNA的目标产生的phasirnas都可以在Begomovirus之间的相互作用中发挥作用N. Benthamiana..
结论
在基因组中发现了总共196个保守的miRNA和197个新的miRNAN. Benthamiana..其中,有几种表现出不同的表达模式N. Benthamiana.植物共感染TYLCCNV和TYLCCNB,以及TYLCCNV和一个突变的TYLCCNB,提示这些mirna在与begomovirus相关的betassatellite和N. Benthamiana.植物。此外,我们的研究表明,这些差异表达的mirna中有几个是phasirna产生的潜在触发器。
方法
小RNA和基因组数据
三组小RNA群体(登录号:GSE26368)N. Benthamiana.是在我们之前的研究中使用高通量测序技术生成的[30.].简而言之,N. Benthamiana.以番茄黄曲叶病毒(TYLCCNV)单独(P1)、TYLCCNV及其相关的betassatellite (TYLCCNB)共感染(P2)或突变TYLCCNB (P3)的植株叶片为材料,构建sRNA cDNA文库。采用高通量测序技术对sRNA文库进行测序。草拟基因组序列N. Benthamiana.下载了http://solgenomics.net(Niben.genome.v0.4.4) [31].
注释miRNA和基因座生成phasirnas
所有小型RNA数据都是通过一套Perl脚本处理的。清洁读数被映射到N. Benthamiana.基因组序列。读取映射到众所周心的非编码RNA(例如,RRNA,TRNA,SNRNA,SNARNA)以进行进一步分析。我们以前的生物信息管道遵循Meyers等人的标准。[35]应用于miRNA注释[36].通过维也纳RNA包进行茎环结构的预测[37].候选人(前miRNA)具有理想的发夹结构,其中包含能够形成miRNA :: miRNA的一对小RNA*选择了小于4个不匹配和2-NT的3'突出的双工。通过搜索编码区域来预测miRNA的目标N. Benthamiana.基因组使用psRNATarget (http://plantgrn.noble.org/psRNATarget/)[38]默认设置(最大预期:3.0;互补得分的长度:20 BP;目标可访问性 - 允许最大能量卸载目标站点:25.0;目标网站周围的侧翼长度为目标可访问性分析:上游17 BP和13 BP在下游;中央不匹配范围导致平移抑制:9-11 nt)。pha利用该算法进行轨迹识别
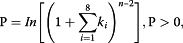
(n = number of phase cycle positions occupied by at least one small RNA read within an eight-cycle window, andk = the total number of reads for all small RNAs with consolidated start coordinates in a given phase within an eight-cycle window) described by Howell et al. [17.我们之前的研究[19.,32].咬合的mirnaspha产生分阶段的小RNA读数的区域内的基因座或它们的侧翼200-NT区域被认为是推定的阶段触发器。在这些pha在基因座中,假定的miRNA剪切位点(在预测miRNA靶位点的10和11位之间)总是在一个相寄存器对应的位置发现。
MiRNA的北印迹分析
从全身感染中提取总RNAN. Benthamiana.留在制造商的指示(Invitrogen,Carlsbad,CA)之后使用Trizol Reagents。总共15-30μg的低分子量(LMW)RNA,由聚乙二醇分离(分子量8000,PEG8000)和NaCl [39],在变性15%聚丙烯酰胺的变性凝胶上分开并转移到杂合+膜。通过UV辐射交联后,用生物素标记的DNA探针杂交以互补于42℃的生物素标记的DNA探针互补过夜。在42℃下在42℃下使用1×SSC和0.5%十二烷基硫酸钠(SDS)进行两次杂交洗涤。根据制造商的说明,用补充在化学发光核酸检测模块(Thermo,Pierce)中的稳定的链霉抗生物素蛋白 - 辣根过氧化物酶缀合物检测杂交信号。为了确认等同的凝胶装载,选择U6作为内部控制,并且斑点也用与U6互补的DNA探针杂交。
miRNA目标的实时量化
使用AMV逆转录酶(Takara,中国大连)按照制造商的说明对1 μg经DNase i处理的总RNA进行逆转录。采用SYBR Green PCR supermix(罗氏,曼海姆,德国)在Lightcycler®real-time PCR系统(罗氏)中进行实时定量PCR,如所述[30.].进行了三种独立的生物学复制,使用用于内部控制的CT值来标准化miRNA靶基因的相对表达GADPH..使用由制造商(Roche)补充的Lighcycler®480软件进行分析。
缩写
- miRNA:
-
microRNA.
- 核:
-
短暂干扰RNA
- phasirna:
-
分阶段,次级siRNA
- 国家统计局:
-
核苷酸结合位点
- tasiRNA:
-
Trans-acting核。
参考
- 1。
Bartel DP:MicroRNAS:基因组学,生物生成,机制和功能。细胞2004年,116.(2): 281 - 297。10.1016 / s0092 - 8674 (04) 00045 - 5
- 2.
kirawesh B,朱家杰,朱军:miRNA和siRNA在生物和非生物应激反应中的作用。Biochim Biophys学报2012年,1819(2):137-148。10.1016 / J.BBBAGRM.2011.05.001
- 3.
Amin I, Patil BL, Briddon RW, Mansoor S, Fauquet CM:一套常见的发育麦芽系统是上调的尼古利亚娜·宾夕法尼亚州通过不同的原野毒病毒。virol j.2011年,8:143.101186 / 1743-422x-8-143
- 4.
Bazzini AA, Hopp HE, Beachy RN, Asurmendi S:烟草花叶病毒蛋白质的侵染和共积累改变了microRNA水平,与症状和植物发育相关。Proc Natl Acad Sci USA2007年,104.(29):12157-12162。10.1073 / PNAS.0705114104
- 5.
郭一,拿jy,袁亚:沿RNA沉默途径的宿主和病毒之间的结构见解拟南芥蒂利亚纳.杂志牧师2013. DOI:10.1111 / BRV.12057
- 6.
何晓芳,方洋洋,冯丽,郭海生:求保守和新微大RNA的表征及其靶标,包括肿瘤诱导的TIR-NBS-LRR类R基因衍生的新型miRNA芸苔属植物.Febs Lett.2008年,582.(16):2445-2452。10.1016 / J.Febslet.2008.06.011
- 7.
Kasschau KD, Xie Z, Allen E, Llave C, Chapman EJ, Krizan KA, CarringtonP1/HC-Pro是一种RNA沉默的病毒抑制因子,可以干扰拟南芥的发育和miRNA功能。Dev细胞2003年,4(2):205-217。10.1016 / s1534-5807(03)00025-x
- 8.
Naqvi AR, Haq QM, Mukherjee SK:对番茄曲叶病毒(tolcndv)侵染番茄叶片的MicroRNA分析表明,mir159/319和mir172的缺失可能与曲叶病有关。virol j.2010,7:281.10.1186 x / 1743 - 422 - 7 - 281
- 9.
Navarro L, Jay F, Nomura K, He SY, Voinnet O:细菌效应蛋白抑制微润松途径。科学2008年,321.(5891):964-967。10.1126 / Science.1159505.
- 10。
Pacheco R,Garcia-Marcos A,Barajas D,Martianez J,Tenllado F:PVX-potyvirus协同感染差异地改变MicroRNA积累尼古利亚娜·宾夕法尼亚州.病毒Res2012年,165(2): 231 - 235。10.1016 / j.virusres.2012.02.012
- 11.
Katiyar-Agarwal S,Gao S,Vivian-Smith A,Jin H:拟南芥中一类新细菌诱导的小rna。基因开发2007年,21(23): 3123 - 3134。10.1101 / gad.1595107
- 12.
du p,吴j,张俊,赵某,郑h,高g,魏l,李y:病毒感染诱导保守的细胞microRNA前体中新型阶段性microRNA的表达。PLOS PAROGOG.2011年,7(8): e1002176。10.1371 / journal.ppat.1002176
- 13.
Allen E, Xie Z, Gustafson AM, Carrington JC:植物中杂交siRNA生物发生期间的MicroRNA定向阶段。细胞2005年,121.(2):207-221。10.1016 / J.Cell.2005.04.004
- 14.
沉D,王某,陈H,朱QH,Helliwell C,风扇L:草族MiR390引导式转型siRNA基因(TAS3)的分子系统发生。植物系统另一个星球2009,283(1-2):125-132。
- 15.
翟杰,济东·德赫,德宝利e,公园,罗森bd,李玉,冈萨雷斯阿,燕z,kitto sl,grusak ma,jackson sa,stacey g,cook博士,绿色pj,谢尔德dj,meyers bc:MicroRNAS作为植物NB-LRR防御基因系列的母稳压器,通过生产分阶段,转作siRNA。基因开发2011年,25(23):2540-2553。10.1101 / gad.177527.111
- 16.
Heisel Se,张Y,Allen E,Guo L,Reynolds TL,Yang X,Kovalic D,Roberts JK:稻粒独特小RNA种群的特征。普罗斯一体2008年,3.(8):E2871。10.1371 / journal.pone.0002871.
- 17.
Howell MD, Fahlgren N, Chapman EJ, Cumbie JS, Sullivan CM, Givan SA, Kasschau KD, Carrington JC:拟南芥中RNA依赖性RNA聚合物酶6 / Dicer-Lige4途径的基因组分析显示依赖于miRNA和染蛋白指导的靶向。植物细胞2007年,19.(3):926-942。10.1105 / TPC.107.050062
- 18.
Johnson C,Kasprzewska A,Tennessen K,Fernandes J,Nan Gl,Walbot V,Sundaresan V,Vance V,Bowman LH:水稻花序发育过程中阶段性小rna的成簇和超簇。基因组Res2009,19.(8): 1429 - 1440。10.1101 / gr.089854.108
- 19.
刘艳,王艳,朱庆华,范丽:野生稻中Phasirnas的鉴定(选用高产稻).植物信号Behav2013年,8(8): e25079。10.4161 / psb.25079
- 20。
Li F, Pignatta D, Bendix C, Brunkard JO, Cohn MM, Tung J, Sun H, Kumar P, Baker B:MicroRNA调节植物生先生免疫受体。美国国立科学院科学研究所2012年,109.(5): 1790 - 1795。10.1073 / pnas.1118282109
- 21。
Shivaprasad PV, Chen HM, Patel K, Bond DM, Santos BA, Baulcombe DC:microRNA超家族调控核苷酸结合位点-富亮氨酸重复序列和其他mrna。植物细胞2012年,24(3): 859 - 874。10.1105 / tpc.111.095380
- 22。
Wang Y, Itaya A, Zhong X, Wu Y, Zhang J, van der Knaap E, Olmstead R, Qi Y, Ding B:调节CA的MicroRNA的功能和演变2+- atp酶,并在番茄生殖生长过程中触发阶段性小干扰rna的形成。植物细胞2011年,23(9): 3185 - 3203。10.1105 / tpc.111.088013
- 23。
Xia R, Zhu H, An YQ, Beers EP, Liu Z:Apple Mirnas和Tasirnas与新型监管网络。基因组Biol.2012年,13.(6): R47。10.1186 / gb - 2012 - 13 - 6 - r47
- 24。
Fei Q, Xia R, Meyers BC:在后谱监管网络中相位,中,小干扰RNA。植物细胞2013年,25(7):2400-2415。10.1105 / TPC.113.114652
- 25.
周X:理解Begomovirus卫星的进步。Annu Rev phytopathol.2013年,51:357-381。10.1146 / annurev-phyto-082712-102234
- 26.
崔X,李g,王d,胡d,周x:一种Begomovirus DNAbeta编码蛋白结合DNA,作为RNA沉默抑制因子,并以细胞核为靶点。J Virol.2005年,79(16):10764-10775。10.1128 / JVI.79.16.10764-10775.2005
- 27.
Cui X,Tao X,谢Y,Fauquet Cm,周X:症状诱导需要与番茄黄叶卷曲卷曲中国病毒有关的DNABETA。J Virol.2004年,78(24): 13966 - 13974。10.1128 / jvi.78.24.13966 - 13974.2004
- 28.
杨杰,iwasaki m,machida c,machida y,zhou x,chua nh:Betac1,Tylccnv的致病性因子与AS1相互作用,以改变叶片发育和抑制选择性茉莉酸反应。基因开发2008年,22(18):2564-2577。10.1101 / gad.1682208
- 29.
杨X,谢y,拉贾p,李思,狼jn,沉q,bisaro dm,周x:抑制β-甲状腺素感染β-SaHH蛋白相互作用的甲基化介导的转录基因沉默。PLOS PAROGOG.2011年,7(10): e1002329。10.1371 / journal.ppat.1002329
- 30。
杨旭,王勇,郭伟,谢勇,谢强,范玲,周旭:深度测序源自精华虫/β外晶体复合物的小干扰RNA的表征。普罗斯一体2011年,6(2): e16928。10.1371 / journal.pone.0016928
- 31。
Bombarely A,Rosli Hg,Vrebalov J,Moffett P,Mueller La,Martin GB:基因组序列草案尼古利亚娜·宾夕法尼亚州加强植物微生物分子生物学研究。Mol植物微生物相互作用2012年,25(12): 1523 - 1530。10.1094 / mpmi - 06 - 12 - 0148 - ta
- 32。
唐S,王y,李z,gui y,小b,谢j,zhu qh,风扇l:烟草伤害和顶部敏感小RNA的识别(尼科尼亚塔哈瓦姆).BMC植物杂志2012年,12:28.10.1186 / 1471-2229-12-28
- 33。
Windels D,Vazquez F:miR393:生长素信号中环境线索的整合者?植物信号Behav2011年,6(11): 1672 - 1675。10.4161 / psb.6.11.17900
- 34。
朱Q,粉丝L,Liu Y,Xu H,Llewellyn D,Wilson I:棉花真菌病原体感染期间NBS-LRR防御基因的miR482调节。《公共科学图书馆•综合》2013年,8(12):E84390。10.1371 / journal.pone.0084390
- 35。
Meyers BC, Axtell MJ, Bartel B, Bartel DP, Baulcombe D, Bowman JL, Cao X, Carrington JC, Chen X, Green PJ, Griffiths-Jones S, Jacobsen SE, Mallory AC, Martienssen RA, Poethig RS, Qi Y, Vaucheret H, Voinnet O, Watanabe Y, Weigel D, Zhu JK:植物microrna注释标准。植物细胞2008年,20.(12):3186-3190。10.1105 / TPC.108.064311
- 36。
王y,白x,yan c,gui y,wei x,zhu qh,guo l,风扇l:野生稻中小rnas的基因组解剖(选用高产稻):米驯化的课程。新的Phytol.2012年,196(3):914-925。10.1111 / J.1469-8137.2012.04304.x
- 37.
Hofacker I, Fontana W, Stadler P, Bonhoeffer L, Tacker M, Schuster P:RNA二级结构的快速折叠和比较。Monatsh化学1994年,125.(2):167-188。10.1007 / bf00818163
- 38.
戴X,赵px:psRNATarget:植物小RNA目标分析服务器。核酸res.2011年,39(Web服务器问题):w155 - 159。
- 39.
Mette MF, Aufsatz W, van der Winden J, Matzke MA, Matzke AJ:通过双链RNA引发的转录沉默和促进剂甲基化。Embo J.2000年,19.(19):5194-5201。10.1093 / emboj / 19.19.5194
确认
我们感谢钱浩浩博士(Csiro植物行业,澳大利亚)的批判读书。该工作得到了中国国家自然科学基金(31200117)和云南省烟草公司(08A05 / 2011YN04)的支持。
作者信息
隶属关系
相应的作者
附加信息
相互竞争的利益
两位作者宣称他们没有相互竞争的利益。
作者的贡献
BX和LF构思了这项研究。XY对miRNA和RT-QPCR分析进行了实验验证。C-Y Y,Y1,Cy和YW鉴定的miRNA和phasirnas并进行了表达分析。LF,BX,XL和YL讨论了数据。LF,XY和C-Y Y写了稿件。所有作者阅读并认可的终稿。
电子辅料
12870 _2013_1484_moesm1_esm.docx
附加文件1:表S1:小rna的统计,可以映射到N. Benthamiana.基因组。的N. Benthamiana.单独用TylCCNV系统感染的植物(P1),或与β卫星TylCCNB(P)或用突变TylCCNB(P3)一起进行RNA提取和小RNA测序。(docx 17 kb)
12870 _2013_1484_moesm5_esm.xlsx
附加文件5:表S5:在N. Benthamiana.在Tylcnv感情之后。(XLSX 18 KB)
权利和权限
本文由BioMed Central Ltd授权发表。这是一篇基于知识共享署名许可协议(http://creativeCommons.org/licenses/by/2.0.),允许在任何媒介上不受限制地使用、分发和复制,但须注明原作的出处。创作共用及公共领域专用豁免书(http://creativecommons.org/publicdomain/zero/1.0/)除非另有说明,否则适用于本文中提供的数据。
关于这篇文章
引用这篇文章
小,B.,阳,X.,YE,CY。等等。一种多样化的miRNA,敏锐地对Begomovirus相关的甜菜丝毛素尼古利亚娜·宾夕法尼亚州.BMC植物杂志14日,60(2014)。https://doi.org/10.1186/1471-2229-14-60
已收到:
接受:
发表:
关键词
- 尼古利亚娜·宾夕法尼亚州
- microrna的
- phasirna.
- Begomovirus.