
- 研究文章gydF4y2Ba
- 开放获取gydF4y2Ba
- 发表:gydF4y2Ba
利用iTRAQ对玉米籽粒发育进行蛋白质组学分析,揭示了不同代谢过程的时间程序gydF4y2Ba
BMC植物生物学gydF4y2Ba体积gydF4y2Ba16gydF4y2Ba文章编号:gydF4y2Ba241gydF4y2Ba(gydF4y2Ba2016gydF4y2Ba)gydF4y2Ba
摘要gydF4y2Ba
背景gydF4y2Ba
玉米的籽粒发育是植物生命周期的一个重要过程,对动物和人类利用玉米作为作物至关重要。然而,人们对控制谷物发育的蛋白质调控网络知之甚少。本研究利用等压标签相对与绝对定量技术(iTRAQ)分析了玉米籽粒发育过程中蛋白质表达的时间变化。gydF4y2Ba
结果gydF4y2Ba
利用基于itraq的蛋白质组学方法,研究了授粉后3 ~ 50 d的玉米籽粒蛋白质及8个发育阶段蛋白质表达的变化。总共鉴定出4751个蛋白质;其中2639个被定量,1235个在不同发育阶段表达水平变化至少1.5倍,被鉴定为差异表达蛋白(DEPs)。DEPs参与不同的细胞和代谢过程,并优先分布于蛋白质合成/目的地和代谢类别。K-means聚类分析显示,在不同发育阶段,不同功能类别/亚类别的蛋白表达是协调的。gydF4y2Ba
结论gydF4y2Ba
结果表明,发育中的玉米籽粒在不同的发育阶段表现出不同的蛋白质组学特征,如早期大量表达与细胞生长/分裂有关的DEPs,而淀粉合成和防御/胁迫相关的DEPs则分别在中后期积累。我们还观察到抗氧化系统中多个蛋白的协同表达,这些蛋白对维持籽粒发育过程中活性氧(ROS)稳态至关重要。特别是一些DEPs,如锌金属硫蛋白II类、丙酮酸正磷酸盐二激酶(PPDK)和14-3-3蛋白,在特定的发育阶段表达发生重大变化,表明它们在玉米籽粒发育中起着重要作用。这些结果为在全球范围内分析蛋白质功能提供了宝贵的资源,也为研究控制粮食产量和品质的潜在蛋白质调控网络提供了新的见解。gydF4y2Ba
背景gydF4y2Ba
谷类作物的谷粒具有较高的农艺价值;玉米尤其如此(gydF4y2Ba玉米gydF4y2BaL.),在世界范围内种植,是最重要的粮食来源,动物饲料和可再生资源的作物之一。提高粮食产量和品质是玉米育种的主要目标。分子生物学技术和基因组学在玉米育种中受到越来越多的关注,因为它们提供了新的和更有效的选择标准[gydF4y2Ba1gydF4y2Ba].因此,更好地理解与籽粒发育相关的代谢过程和潜在分子机制,将为未来提高籽粒产量和品质提供新的认识。gydF4y2Ba
在过去的几十年里,对玉米籽粒发育的认识取得了很大的进展,玉米籽粒发育由双施肥过程开始,并分为三个主要阶段:滞后期、灌浆期和成熟期[gydF4y2Ba2gydF4y2Ba,gydF4y2Ba3.gydF4y2Ba].授粉后12天(DAP)为迟滞期,其特征是细胞数量和大小的快速增长;这种增加决定了储存分子随后积累的汇的大小。灌浆期为12 - 40dap,以贮存分子的合成和积累为特征。在这一阶段,以进口蔗糖为原料合成的淀粉为主要贮藏成分,由直链淀粉和支链淀粉组成。各种酶合成淀粉,然后在淀粉质体中将分子修剪和包装成半结晶淀粉颗粒[gydF4y2Ba4gydF4y2Ba- - - - - -gydF4y2Ba6gydF4y2Ba].成熟期为40 - 70 DAP,以颗粒脱水为特征,并逐渐进入休眠状态。每个阶段的持续时间取决于遗传背景、环境和文化条件[gydF4y2Ba7gydF4y2Ba].虽然我们对籽粒发育过程中形态和生理变化的了解已经增加,但其潜在的分子调控机制仍在很大程度上是未知的[gydF4y2Ba8gydF4y2Ba- - - - - -gydF4y2Ba10gydF4y2Ba].gydF4y2Ba
基因活性和功能的鉴定是探索分子调控机制的有效方法。利用微阵列、cDNA文库和RNA测序(RNA-seq)进行的大规模全基因组表达分析已经描述了大量在玉米籽粒发育过程中胚胎发生或储存化合物积累中优先表达的基因[gydF4y2Ba1gydF4y2Ba,gydF4y2Ba9gydF4y2Ba- - - - - -gydF4y2Ba13gydF4y2Ba].例如,利用RNA-seq策略对玉米胚、胚乳和全粒从受精到成熟的过程进行了动态转录组学分析,发现了26105个与粒发育程序相关的基因;此外,这些基因中有1258个被确定为籽粒特异性基因[gydF4y2Ba10gydF4y2Ba].虽然已经报道了与籽粒发育有关的基因的信息,但在蛋白质水平上缺乏同等的细节,尽管它们是细胞活动的直接调节者。更重要的是,转录模式并不总是与相应蛋白的表达直接相关,就像在玉米中所显示的那样[gydF4y2Ba14gydF4y2Ba)、大米(gydF4y2Ba15gydF4y2Ba)、棉花(gydF4y2Ba16gydF4y2Ba),而gydF4y2Ba拟南芥gydF4y2Ba[gydF4y2Ba17gydF4y2Ba].因此,直接的蛋白质组学研究对监测籽粒发育状况也至关重要。gydF4y2Ba
迄今为止,关于籽粒发育的蛋白质组学研究主要采用双向凝胶电泳(2-DE)。这样的研究已经在许多物种中进行,包括水稻[gydF4y2Ba15gydF4y2Ba,gydF4y2Ba18gydF4y2Ba,gydF4y2Ba19gydF4y2Ba)、小麦(gydF4y2Ba20.gydF4y2Ba),gydF4y2Ba拟南芥gydF4y2Ba[gydF4y2Ba17gydF4y2Ba],大麦[gydF4y2Ba21gydF4y2Ba], castor [gydF4y2Ba22gydF4y2Ba),gydF4y2BaMedicago truncatulagydF4y2Ba[gydF4y2Ba23gydF4y2Ba,大豆[gydF4y2Ba24gydF4y2Ba,gydF4y2Ba25gydF4y2Ba].然而,由于2-DE存在无法分离疏水蛋白、识别率低、缺乏准确的定量信息等固有限制,某些类型的蛋白质无法通过2-DE进行分析[gydF4y2Ba26gydF4y2Ba,gydF4y2Ba27gydF4y2Ba].最近,利用等压标签进行相对和绝对定量(iTRAQ)作为一种基于质谱的定量技术已被开发出来;这项技术克服了2-DE的一些局限性,特别是对于多个样品,并允许鉴定更多的蛋白质,以提供更可靠的定量信息[gydF4y2Ba28gydF4y2Ba,gydF4y2Ba29gydF4y2Ba].iTRAQ技术的优势已经被用来鉴定和量化水稻胚胎发育中的2165个蛋白质[gydF4y2Ba30.gydF4y2Ba和小麦籽粒中的1815种蛋白质[gydF4y2Ba31gydF4y2Ba].gydF4y2Ba
在过去,对玉米全粒或胚、胚乳进行了多种蛋白质组学分析。基于2-DE, Méchin等[gydF4y2Ba32gydF4y2Ba]建立了玉米胚乳蛋白质组参考图谱,共鉴定出504个蛋白质,主要属于代谢和蛋白质目的类。随后,他们在4 - 40 DAP的7个发育阶段对409种蛋白质进行了定量分析,表明这些蛋白质的动态表达模式与从细胞生长和分化到存储的重要发育转变相一致[gydF4y2Ba8gydF4y2Ba].为了探索玉米籽粒灌浆的关键调控因子,Jin等[gydF4y2Ba7gydF4y2Ba]通过2-DE分析发现,在3个优质玉米杂交种线性灌浆期(17-28 DAP),胚乳中有39个蛋白和43个蛋白表达差异,进一步的功能分析表明,胚乳中与糖酵解和氧化还原稳态有关的蛋白被强调,而胚乳中与脂肪酸生物合成有关的蛋白被强调。通过2-DE对玉米籽粒进行老化处理后,发现有40个蛋白表达存在差异,说明人工老化影响了干玉米籽粒的蛋白质组[gydF4y2Ba33gydF4y2Ba].在其他使用2-DE的研究中,研究了玉米胚脱水耐性相关蛋白的表达水平[gydF4y2Ba34gydF4y2Ba[]并对籽粒活力进行了调查[gydF4y2Ba35gydF4y2Ba].然而,由于2-DE法的局限性,这些研究只能研究相对较少的蛋白质。在最近的一项研究中,Walley等人利用质谱技术[gydF4y2Ba14gydF4y2Ba]根据蛋白质丰度和磷酸化水平绘制了发育中的玉米籽粒的蛋白质型图谱。结果,在不同的粒室和发育阶段,包括胚胎(吸胀后20、38个DAP和2天)、胚乳(8、10、12和27个DAP)和粘粉层/果皮(27个DAP),定量检测了14165个蛋白质和18405个磷酸肽(来自4511个蛋白质),该研究进一步揭示了许多最丰富的蛋白质与可检测的mrna水平无关[gydF4y2Ba14gydF4y2Ba].该图谱为识别与粮食发育和储粮产品生产相关的激酶-底物关系以及重建支撑粮食发育和储粮产品生产的生化和信号网络提供了丰富的资源。尽管玉米籽粒蛋白质组学研究已经开展了大量的工作,但这些研究主要集中在玉米籽粒不同成分(胚、胚乳和糊粉/果皮)和几个时间点上。与此同时,玉米的基因组更大,蛋白质组更复杂gydF4y2Ba拟南芥gydF4y2Ba对于水稻而言,玉米籽粒发育的调控机制仍需进一步研究。重要的是,据我们所知,基于iTRAQ对玉米籽粒整个发育过程进行系统的蛋白质组学分析尚未见报道。因此,我们利用iTRAQ技术分析了3 - 50 DAP (3 - 50 DAP)三个主要发育过程中玉米籽粒蛋白质表达的动态变化。我们的研究结果揭示了与籽粒发育相关的蛋白质表达模式的全球变化,为在全球范围内分析蛋白质功能提供了宝贵的资源,并为研究控制籽粒产量和品质的潜在蛋白质调控网络提供了新的见解。gydF4y2Ba
结果与讨论gydF4y2Ba
玉米籽粒八个发育阶段的生理特性gydF4y2Ba
在3、5、10、15、20、30、40和50 DAP取样全玉米籽粒。gydF4y2Ba1gydF4y2Ba),并在每个采样日记录发育颗粒的特征(图。gydF4y2Ba1 bgydF4y2Ba- - - - - -gydF4y2BadgydF4y2Ba).在籽粒发育过程中,鲜、干粒重均从3 - 10 DAP缓慢增加,随后增加更快,达到40 DAP(图;gydF4y2Ba1 bgydF4y2Ba).40 DAP后,干重持续增加,直到50 DAP,鲜重下降,表明40 DAP后发育的籽粒进入干燥阶段。总淀粉含量在10 - 30 DAP之间快速增加,然后缓慢增加,直到50 DAP(图。gydF4y2Ba1 cgydF4y2Ba表明10 ~ 30dap是籽粒淀粉合成积累的关键阶段。胚乳细胞增殖分析显示,胚乳细胞数量从3 DAP迅速增加到10 DAP,然后缓慢增加,在20 DAP达到最大值(图2)。gydF4y2Ba1 dgydF4y2Ba).结果表明,在10个DAP发育的籽粒中,细胞主要进行活跃分裂和分化,40 DAP后籽粒灌浆,40 DAP后进入干燥阶段。因此,这三个时期大致代表了玉米籽粒发育的三个主要阶段,即早期(3-10 DAP)、中期(10-40 DAP)和晚期(40-50 DAP)。gydF4y2Ba

试验期间玉米籽粒发育情况。gydF4y2Ba一个gydF4y2Ba玉米籽粒发育的八个阶段。gydF4y2BabgydF4y2Ba发育中籽粒鲜、干重的变化。每个阶段至少分析100粒。gydF4y2BacgydF4y2Ba发育中籽粒总淀粉含量的变化。数值表示为谷物干重的百分比。gydF4y2BadgydF4y2Ba籽粒胚乳细胞数量的动态变化。误差条表示三次重复的标准差gydF4y2Ba
玉米籽粒蛋白质的鉴定与相对定量gydF4y2Ba
使用基于itraq的蛋白质组学分析总蛋白和蛋白表达变化,3个生物重复。还采用了严格的鉴别和质量标准(详见“方法”)。分析发现玉米籽粒中有4751个蛋白,其中2755个蛋白是在两次生物重复中常见的(图2)。gydF4y2Ba2gydF4y2Ba;额外的文件gydF4y2Ba1gydF4y2Ba);其中2639个被定量(附加文件gydF4y2Ba2gydF4y2Ba).鉴定和定量的蛋白质数量比之前使用2-DE进行的蛋白质组学分析更多[gydF4y2Ba8gydF4y2Ba,gydF4y2Ba32gydF4y2Ba],这清楚地表明,与传统的凝胶法相比,iTRAQ技术在蛋白质鉴定和定量方面具有更大的潜力。同时,与最近的蛋白质组学分析相比,该分析通过质谱法定量了正在发育的玉米胚(吸胀后20、38个DAP和2天)、胚乳(8、10、12和27个DAP)和糊粉/果皮(27个DAP)中的14,165个蛋白质和18,405个磷酸肽(来自4511个蛋白质)[gydF4y2Ba14gydF4y2Ba],在我们的研究中,鉴定和定量的蛋白数量仍然相对较少。然而,本研究描述了玉米籽粒蛋白质组在整个发育阶段的动态变化。利用发育过程中1.5倍的表达截断变化(p≤0.05)来确定差异表达蛋白(DEPs)丰度的显著变化。共有1235个蛋白被归类为DEPs, K-means聚类分析将这些蛋白划分为5个表达聚类组(c0, c1, c2, c3和c4;表格gydF4y2Ba1gydF4y2Ba;无花果。gydF4y2Ba3.gydF4y2Ba).最大的簇为c0,该簇中含有466个蛋白质;从3到50 DAP,这些蛋白的表达逐渐下降。其次是c2(279个蛋白)和c1(243个蛋白)。c2蛋白表达量在30 DAP时逐渐增加,在50 DAP时达到最大值。与此相反,c1蛋白在15-20 DAP时大量积累,偶尔在30 DAP时积累,此后逐渐减少。Cluster c3包含82个蛋白,它们的表达模式与c2相似,只是在30到50 DAP之间有较大的增长。簇c4由165个蛋白组成,与c1蛋白相比,其表达高峰分别在6和50 DAP。这些结果表明,不同的蛋白质调控模式与籽粒发育的早、中、后期有关。gydF4y2Ba

表示已鉴定的蛋白质在三个生物重复序列中重叠的维恩图。Bio1, Bio2, Bio3分别代表第一个,第二个和第三个生物复制gydF4y2Ba

8个发育阶段功能性DEPs的K-means聚类分析。功能dep列在附加文件中gydF4y2Ba4gydF4y2Ba,以及它们的集群分配信息gydF4y2Ba
在1235个DEPs中,572个被标注为未鉴定蛋白。为了获得这些蛋白的功能信息,我们进行了BLAST分析,以寻找同源蛋白;在437个未鉴定的蛋白序列中,发现了其他物种的同源序列(附加文件gydF4y2Ba3.gydF4y2Ba).根据UniProt中假定的生物学功能和玉米胚乳功能分类方案[gydF4y2Ba8gydF4y2Ba和米粒[gydF4y2Ba15gydF4y2Ba, 1235个DEPs被划分为不同的功能类别。参与蛋白质合成/目标和代谢的蛋白质组成了最大的类群,分别约占25.18和20%(图。gydF4y2Ba4gydF4y2Ba),表明在籽粒发育过程中代谢和蛋白质合成/归宿的功能重要性。为了获得关于这两个功能类别的更详细的信息,涉及蛋白质合成/目标和代谢的DEPs分别被划分为5和11个子类别(表gydF4y2Ba1gydF4y2Ba).gydF4y2Ba

玉米籽粒发育过程中鉴定的DEPs的功能分布gydF4y2Ba
籽粒发育迟滞期蛋白质表达特征gydF4y2Ba
0-10 DAP的籽粒滞后期表现为细胞分裂活跃,增大籽粒库尺寸,为后续贮藏物质的积累提供条件。先前的蛋白质组学分析表明,细胞骨架蛋白(肌动蛋白和微管蛋白)在胚胎发生过程中对细胞分裂和增大起着至关重要的作用[gydF4y2Ba36gydF4y2Ba,在这个阶段积累到了最高水平。在本研究中,我们证实了大多数与细胞生长/分裂相关的DEPs(113个蛋白中的75个)优先积累于这一阶段(c0;表格gydF4y2Ba1gydF4y2Ba),不仅包括细胞骨架蛋白,还包括其他蛋白,如增殖细胞核抗原和组蛋白(附加文件gydF4y2Ba4gydF4y2Ba).增殖细胞核抗原参与DNA修复和细胞周期调控[gydF4y2Ba37gydF4y2Ba].同时,细胞壁和单位膜(细胞的结构成分)的形成也得到了加强[gydF4y2Ba38gydF4y2Ba,gydF4y2Ba39gydF4y2Ba].因此,大多数与细胞壁形成相关的DEPs(25个蛋白中的15个),如udp -葡萄糖6-脱氢酶(B6T9P0和B6TBY8)和木葡聚糖内转糖基化酶/水解酶(B4G1Z2)在这一早期表现出最大的积累(附加文件)gydF4y2Ba4gydF4y2Ba).udp -葡萄糖6-脱氢酶参与细胞壁多糖的合成,而木葡聚糖内转糖苷酶/水解酶的作用是使细胞壁松动和延长[gydF4y2Ba38gydF4y2Ba].在细胞膜的形成过程中,大约一半的脂质/固醇代谢相关蛋白(50个蛋白中的23个)积累增强,与细胞壁相关蛋白(c0;表格gydF4y2Ba1gydF4y2Ba);脂质/甾醇代谢相关蛋白中有皂苷样B型蛋白(B6T780),它可以与脂质相互作用[gydF4y2Ba40gydF4y2Ba并可作为表面活性剂,以降低表面张力[gydF4y2Ba41gydF4y2Ba].充分阐明该蛋白在降低玉米籽粒发育早期细胞扩张导致的细胞表面张力方面的作用将是有意义的。gydF4y2Ba
在水稻早期发育过程中发现了活性蛋白周转[gydF4y2Ba15gydF4y2Ba].在这里,我们的分析表明,三分之一的DEPs参与蛋白水解(60个蛋白中的20个)在早期(c0;表格gydF4y2Ba1gydF4y2Ba).其中一些DEPs是泛素/26S蛋白酶体途径(20个蛋白中的9个)的关键组成部分,这是多种细胞和发育事件的重要蛋白质降解途径[gydF4y2Ba42gydF4y2Ba,gydF4y2Ba43gydF4y2Ba].同时,大量蛋白质合成相关蛋白(114个蛋白中56个)和蛋白质运输相关蛋白(39个蛋白中23个)与蛋白质水解相关蛋白(c0;表格gydF4y2Ba1gydF4y2Ba).综上所述,蛋白质周转和重排在玉米早期细胞分裂和增大过程中也起着重要作用。gydF4y2Ba
玉米籽粒发育过程的蛋白质组学分析揭示了糖酵解和三羧酸(TCA)循环在蛋白质水平上的调控[gydF4y2Ba7gydF4y2Ba,gydF4y2Ba8gydF4y2Ba].这两种途径相关蛋白表达量的显著下降标志着籽粒进入成熟期,说明糖酵解和TCA循环的调控对籽粒发育的重要性。然而,戊糖磷酸途径(penttose phosphate pathway, PPP)作为玉米籽粒中另一个重要的呼吸途径,其调控机制却鲜为人知。PPP在植物代谢中起着核心作用,可为多种代谢途径提供还原力和磷酸戊糖[gydF4y2Ba44gydF4y2Ba].还原力由葡萄糖-6-磷酸1-脱氢酶(G6PDH)和6-磷酸葡萄糖酸脱氢酶(6PGDH)在PPP的氧化部分(oxPPP)产生;G6PDH被认为是oxPPP的速率限制[gydF4y2Ba44gydF4y2Ba,gydF4y2Ba45gydF4y2Ba].我们的分析确定了7个与PPP相关的DEPs,包括2个G6PDHs (B6TSB3和C0PFX0)和3个6pdhs (A0A096SF47、Q9SBJ3和B4FSV6);这两种PPP蛋白在之前的玉米籽粒蛋白质组学研究中未被发现[gydF4y2Ba8gydF4y2Ba,gydF4y2Ba34gydF4y2Ba,gydF4y2Ba35gydF4y2Ba].令人惊讶的是,这些PPP蛋白类型在早期优先积累(表gydF4y2Ba1gydF4y2Ba;额外的文件gydF4y2Ba4gydF4y2Ba),表明oxPPP是高度活跃的。研究发现,在蓖麻籽粒发育过程中,G6PDH也表现出类似的表达下降模式[gydF4y2Ba22gydF4y2Ba].综上所述,活性PPP对玉米早期籽粒发育至关重要,可能为这一阶段膜合成和细胞分裂所需的脂肪酸和核酸合成提供了还原力和磷酸戊糖[gydF4y2Ba44gydF4y2Ba].gydF4y2Ba
籽粒灌浆是由细胞生长向淀粉合成积累过渡的过程gydF4y2Ba
在籽粒发育中期(10-40 DAP),籽粒细胞数量和大小略有增加,而贮藏物质(主要是淀粉)开始快速合成和积累。一个引人注目的发现是,与细胞生长相关的DEPs较早期迅速下调,而与淀粉合成相关的蛋白在中期达到最大水平(表)gydF4y2Ba1gydF4y2Ba),反映了从细胞分裂分化到籽粒灌浆的转变。在以往的研究中,玉米籽粒发育过程中淀粉合成相关蛋白表达变化的蛋白质组学分析受到了很多限制,因为这些蛋白在籽粒中检测到少量[gydF4y2Ba7gydF4y2Ba,gydF4y2Ba8gydF4y2Ba这些蛋白质的定量信息不能覆盖整个籽粒发育阶段[gydF4y2Ba14gydF4y2Ba].相反,iTRAQ方法鉴定和量化了大量与淀粉生物合成相关的关键蛋白,包括蔗糖合酶(SuSy)、adp -葡萄糖焦磷酸化酶(AGPase)、adp -葡萄糖脆-1转运蛋白(BT1)、淀粉合酶(SS)、淀粉分支酶IIb (SBEIIb)、异淀粉酶I (ISAI)和淀粉磷酸化酶(SP)。这些蛋白大部分被归为c1聚类(表gydF4y2Ba1gydF4y2Ba);本研究为玉米籽粒发育过程中淀粉合成的研究提供了一个全面的视角。gydF4y2Ba
- 1.gydF4y2Ba
SuSy, AGPase和BT1gydF4y2Ba
在植物库器官中,蔗糖用于淀粉合成的初步动员是由SuSy [gydF4y2Ba46gydF4y2Ba];AGPase催化淀粉合成的第一个关键调控步骤,将葡萄糖-1-磷酸(G1P)转化为adp -葡萄糖(ADP-Glu) [gydF4y2Ba5gydF4y2Ba].据报道,在玉米籽粒中,SuSy转录水平在发育中期之前会增加,之后会下降[gydF4y2Ba10gydF4y2Ba].在我们的研究中,SuSy的4个同工酶被鉴定为DEPs(图。gydF4y2Ba5gydF4y2Ba).其中一个SuSy同工酶(C0P6F8)的表达水平稳定到20dap,此后迅速下降。另外2个同工酶(Q93WS3和B6U1D7)增长到20 DAP, 1个同工酶(K7VDR8)增长到30 DAP,然后全部下降;这些模式与转录的模式一致。这表明两种类型的SuSy可能在籽粒发育早期(I型)和中期(II型)活跃。AGPase的五种亚型被鉴定为DEPs(图。gydF4y2Ba5gydF4y2Ba),其中4种蛋白的表达水平在20 DAP左右达到高峰,此时淀粉合成最活跃。在玉米和水稻等谷物中,细胞质AGPase贡献了大部分的AGPase总活性[gydF4y2Ba5gydF4y2Ba].用于淀粉生物合成的ADP-Glu大部分在胞浆中合成,然后通过BT1输入到淀粉体中,BT1的活性与ADP-Glu的转运效率密切相关[gydF4y2Ba47gydF4y2Ba,gydF4y2Ba48gydF4y2Ba].在我们的研究中发现了1例BT1。gydF4y2Ba5gydF4y2Ba),在20 DAP左右达到峰值,与AGPase表达模式相似。AGPase和BT1的共同表达可以保证淀粉合成所需的ADP-Glu的有效供应。gydF4y2Ba
图5gydF4y2Ba 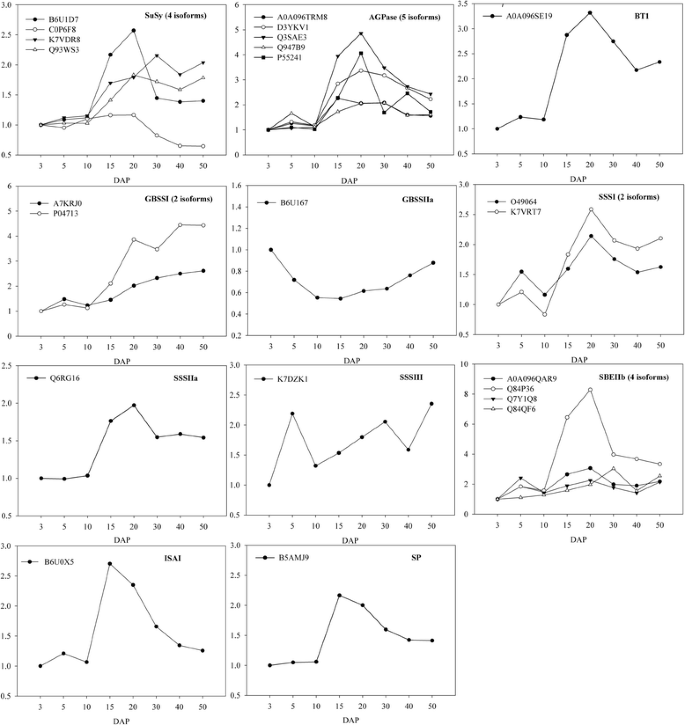
参与淀粉代谢的蛋白表达水平。纵轴表示各发育阶段各亚型与3dap的相对表达比(DAP;水平轴)gydF4y2Ba
- 2.gydF4y2Ba
SS和SBEIIbgydF4y2Ba
SS通过颗粒结合淀粉合成酶(GBSSI和II)和可溶性淀粉合成酶(SSSI到IV)的作用延长葡聚糖链,它们分别负责直链淀粉和支链淀粉的合成[gydF4y2Ba5gydF4y2Ba].我们的研究发现了两种GBSSI亚型的差异表达模式,一种是GBSSIIa亚型,两种是SSSI亚型,一种是SSSIIa亚型,还有一种是SSSIII亚型。gydF4y2Ba5gydF4y2Ba).随着淀粉积累量的增加,GBSSIIa下调,而GBSSI则持续上调。gydF4y2Ba5gydF4y2Ba),支持之前关于GBSSI致力于淀粉酶合成的报道[gydF4y2Ba5gydF4y2Ba,gydF4y2Ba49gydF4y2Ba].以往研究发现,大部分SSS活性依赖于SSSI和SSSIII产物[gydF4y2Ba50gydF4y2Ba].我们发现SSSI在20 DAP左右达到峰值,与淀粉积累的动态变化相一致,而SSSIII丰度在6和50 DAP达到峰值。有趣的是,SSSIIa也被鉴定为DEP,并被发现在20 DAP表现出峰值表达,与SSSI的表达模式相似(图。gydF4y2Ba5gydF4y2Ba).这一结果表明,需要重新评估SSSIIa对玉米籽粒全球SSS活性的贡献。玉米中存在三种类型的淀粉分枝酶(SBEIa, SBEIIa和SBEIIb),它们参与决定支链淀粉的分枝密度和分枝模式[gydF4y2Ba51gydF4y2Ba].SBEIIb的四种亚型被鉴定为DEPs(图。gydF4y2Ba5gydF4y2Ba);其中3种亚型的峰值表达在20dap左右,与AGPase、BT1、SSSI和SSSIIa的表达模式相似。这些结果表明,AGPase、BT1、SBE和SSS的同步表达和/或活性对淀粉合成至关重要。gydF4y2Ba
- 3.gydF4y2Ba
ISA和SPgydF4y2Ba
ISA由ISAI、ISAII和ISAIII三种类型组成,是一种淀粉脱支酶,水解聚葡聚糖的α- 1,6 -葡萄糖键。相反,SP催化葡萄糖基从G1P可逆转移到α- 1,4 -链葡聚糖链的非还原性末端[gydF4y2Ba5gydF4y2Ba].在本研究中,两个DEPs被鉴定为ISAI和SP(图。gydF4y2Ba5gydF4y2Ba),两者都在15 DAP左右达到表达高峰。在水稻籽粒中,ISAs在灌浆期积累最多[gydF4y2Ba15gydF4y2Ba],他们通过影响淀粉颗粒的起始来确定淀粉颗粒的数量[gydF4y2Ba52gydF4y2Ba].在玉米中,正常淀粉颗粒生长也需要ISAs [gydF4y2Ba53gydF4y2Ba].最初,人们认为SP在淀粉代谢中具有降解功能而不是生物合成功能;然而,对水稻籽粒突变体的遗传分析表明,SP在促进淀粉合成起始过程中发挥作用[gydF4y2Ba54gydF4y2Ba].最近的一项研究提出,SP在建立线性短链麦芽低聚糖库方面具有重要作用,可作为水稻淀粉合成的引物[gydF4y2Ba55gydF4y2Ba].因此,在玉米籽粒中,ISA和SP在淀粉快速积累阶段共表达,提示SP可能参与淀粉合成起始过程。gydF4y2Ba

籽粒成熟期蛋白质表达特征gydF4y2Ba
成熟期(40 ~ 50 DAP),籽粒贮藏物质积累缓慢,同时开始脱水,最终获得脱水耐性。几乎所有与储存相关的DEPs(21种蛋白质中的16种;例如,球蛋白、豆科蛋白、α和γ玉米醇溶蛋白)在这一阶段以最大水平积累(c2和c3;表格gydF4y2Ba1gydF4y2Ba).这些蛋白质在萌发过程中逐渐退化,是幼苗生长发育的氮源和碳骨架。初生多肽折叠成成熟蛋白质是由许多分子伴侣和蛋白质折叠催化剂控制的。因此,相当大比例的DEPs参与了蛋白质的折叠/修饰(77种蛋白质中的29种;例如,热休克蛋白和伴侣蛋白(chaperones/chaperonins)与存储蛋白(c2和c3;表格gydF4y2Ba1gydF4y2Ba);这些蛋白是研究玉米籽粒贮藏蛋白折叠组装的理想蛋白。令人惊讶的是,许多与蛋白质水解相关的DEPs(60个蛋白中的23个)在这个阶段也显示出显著的积累(c2和c3;表格gydF4y2Ba1gydF4y2Ba),其中大部分是泛素蛋白(23种蛋白中的13种)。泛素是一种高度保守的蛋白质,泛素化是所有真核生物信号转导的主要修饰因子。泛素/26S蛋白酶体途径是多种细胞和发育事件的主要蛋白质水解机制[gydF4y2Ba42gydF4y2Ba,gydF4y2Ba43gydF4y2Ba].因此,泛素蛋白表达丰度的变化表明泛素化可能在成熟阶段发挥重要作用。gydF4y2Ba
有趣的是,我们还发现其中4种油苷(B6SIZ2、B6SI42、B6TMT0和P21641)和1种甾体油苷(B6UGU4)在后期积累水平最高gydF4y2Ba4gydF4y2Ba).油质脂质和固醇脂质都可以嵌入到包裹在油体周围的单层磷脂膜中,而油体是谷类作物中主要的脂质储存细胞器[gydF4y2Ba56gydF4y2Ba,gydF4y2Ba57gydF4y2Ba].油黏蛋白是阻止油体聚结的主要蛋白质[gydF4y2Ba56gydF4y2Ba]及/或调节油体的大小[gydF4y2Ba57gydF4y2Ba].在玉米籽粒脱水过程中,由于细胞质的压缩,油体可能会合并;因此,油质和固醇油质的积累可能对玉米籽粒的油体结构和脂质积累有一定的控制作用。gydF4y2Ba
在成熟期,与应激/防御相关的DEPs(105个蛋白中76个)大量积累(c2和c3;表格gydF4y2Ba1gydF4y2Ba);例如晚期胚胎发生丰富(LEA)蛋白,包括1个来自LEA组(K7VM99), 3个来自LEA 14-A (B4F9K0, B4G1C1和B6UH99), 3个来自LEA组(B6SID7, B6SJ28和B6UI06), 4个来自LEA D-34 (A0A096TZ44, B6SN63, B6SNS4和B6UH67)gydF4y2Ba4gydF4y2Ba).水稻在籽粒成熟期也观察到LEA蛋白表达增加[gydF4y2Ba19gydF4y2Ba和小麦[gydF4y2Ba31gydF4y2Ba].LEA蛋白的存在和水平的增加与脱水耐受性相关[gydF4y2Ba58gydF4y2Ba,gydF4y2Ba59gydF4y2Ba],它们的表达也会被诱导以响应各种非生物或生物的压力[gydF4y2Ba59gydF4y2Ba].因此,LEA蛋白可能参与保护谷物在发育后期免受严重脱水。最近的一项蛋白质组学研究表明,LEA蛋白可能也是玉米生存能力的一个重要因素[gydF4y2Ba35gydF4y2Ba].综上所述,这些胁迫/防御相关蛋白表明玉米籽粒发育过程中存在一种协同的不良反应和防御机制,可以保护籽粒免受不利环境的影响。更重要的是,与胁迫/防御相关的单个或多个蛋白质可能作为抗性玉米的蛋白质标记。gydF4y2Ba
籽粒发育过程中ROS的稳态调节gydF4y2Ba
在玉米籽粒发育早期(4-14 DAP),氧水平较高,线粒体呼吸强烈;在后期,谷物主要受到脱水、缺氧等各种胁迫[gydF4y2Ba60gydF4y2Ba].在应激条件下,可连续产生活性氧(ROS),如超氧自由基、过氧化氢和羟基自由基[gydF4y2Ba61gydF4y2Ba];这些物质会破坏细胞成分,但对许多生物过程的信号调节也很重要[gydF4y2Ba62gydF4y2Ba].细胞已经发展出广泛的抗氧化系统来维持ROS动态平衡[gydF4y2Ba61gydF4y2Ba,gydF4y2Ba62gydF4y2Ba].在玉米籽粒发育过程中,有58个DEPs被鉴定为ROS相关蛋白(如抗坏血酸脱氢还原酶、谷胱甘肽s转移酶、超氧化物歧化酶和硫氧还蛋白);它们参与了多种抗氧化系统[gydF4y2Ba61gydF4y2Ba].进一步分析表明,14、12和17个DEPs分别在早期、中期和晚期达到最大积累水平。15个DEPs在早期和晚期都显示出显著的聚集(表)gydF4y2Ba1gydF4y2Ba;额外的文件gydF4y2Ba4gydF4y2Ba).这些蛋白的不同表达模式提示了多种复杂的抗氧化系统在不同发育阶段的协同作用。已鉴定的DEPs中有1-cys-过氧化物还蛋白(cys- prx;A2SZW8)和II类锌金属硫蛋白(P43401)均在后期积累量最大(附加文件gydF4y2Ba4gydF4y2Ba).虽然cys- prx被提议参与维持谷物休眠[gydF4y2Ba63gydF4y2Ba,其他一些证据表明,它与休眠没有显著的相关性,但确实具有更高的抗氧化应激能力[gydF4y2Ba64gydF4y2Ba]或可作为传感器,在不利条件下抑制种子萌发[gydF4y2Ba65gydF4y2Ba].最近的一项研究进一步提出,在恶劣条件下,籽粒中cys- prx可能作为分子伴侣保护籽粒发育[gydF4y2Ba66gydF4y2Ba].锌金属硫蛋白II类蛋白属于金属硫蛋白家族,可能在维持必要的金属稳态、有毒金属的解毒和细胞间氧化应激保护中发挥重要作用[gydF4y2Ba67gydF4y2Ba,gydF4y2Ba68gydF4y2Ba].根据这些功能,高水平的II类锌金属硫蛋白积累可能是抵御细胞间氧化应激功能所必需的,并/或提供了一种储存锌和其他金属在萌发后生长所需的一种手段。类似地,在小麦籽粒发育后期也观察到锌金属硫蛋白II类蛋白的积累高峰[gydF4y2Ba69gydF4y2Ba].gydF4y2Ba
PPDK在淀粉合成和能量供应中的可能作用gydF4y2Ba
丙酮酸正磷酸盐二激酶(PPDK)催化丙酮酸(Pyr)、ATP和Pi可逆转化为磷酸烯醇式丙酮酸(PEP)、AMP和PPi。PPDK是C4循环中的一种光合酶,但许多蛋白质组学研究发现,PPDK的多种异构体在水稻等谷物的发育中大量积累[gydF4y2Ba15gydF4y2Ba和小麦[gydF4y2Ba31gydF4y2Ba],表明PPDK在籽粒发育过程中起重要作用。在玉米基因组中,有两个位点编码三种PPDK蛋白:gydF4y2BaPPDKZM1gydF4y2Ba通过交替剪接编码c4型叶绿体PPDK1和胞质PPDK1gydF4y2BaPPDKZM2gydF4y2Ba贡献另一种胞质PPDK2 [gydF4y2Ba70gydF4y2Ba].在本研究中,PPDK1 (B7ZYP6)和PPDK2 (K7UZT6)被鉴定为DEPs,并被归为聚类1,在早期表现为低表达水平,在15-20 DAP显著增加,此后下降(表)gydF4y2Ba1gydF4y2Ba;额外的文件gydF4y2Ba4gydF4y2Ba).而在水稻籽粒中,PPDK蛋白主要表达在灌浆前期,而非灌浆期[gydF4y2Ba71gydF4y2Ba].不同物种间的不同表达模式,PEP与Pyr之间的循环以及PPDK蛋白与atp之间的平衡,可能反映了PPDK蛋白在籽粒发育过程中的多重功能[gydF4y2Ba71gydF4y2Ba].然而,它们在玉米籽粒发育中的确切作用仍不清楚。gydF4y2Ba
基于我们的研究结果,PPDK可能优先作用于PEP到Pyr的形成方向。与PPDK表达模式一致的是,大多数与糖酵解相关的DEPs(24个蛋白中的15个)在15 - 20 DAP (c1;表格gydF4y2Ba1gydF4y2Ba).例外的是丙酮酸激酶(PK, B6TII5),它是一种不可逆的酶,可以将PEP转化为Pyr;该酶在籽粒发育过程中表现为连续下调gydF4y2Ba4gydF4y2Ba).这些结果表明,参与糖酵解活性的蛋白参与了导致PEP产量增加的反应,而PEP由于PK的下调而不能有效地转化为Pyr。同时,大部分参与酒精发酵的蛋白(7个蛋白中的5个),如丙酮酸脱羧酶、乙醇脱氢酶以及参与丙酮酸脱氢酶复合体的蛋白(5个蛋白中的3个)被优先归类为聚1(表1)gydF4y2Ba1gydF4y2Ba;额外的文件gydF4y2Ba4gydF4y2Ba).这种表达模式与PPDK和糖酵解的表达模式相似。这些结果表明,Pyr作为一个反应底物,是活性丙酮酸脱氢酶复合物和酒精发酵途径的主要目标。因此,作为一种补充途径,PPDK可能催化PEP产生足够的Pyr供上述两个过程使用。gydF4y2Ba
重要的是,Pyr的形成可能有利于籽粒灌浆期淀粉的合成和能量的供应。我们的结果显示,包括AGPase在内的大多数淀粉合成相关蛋白被归为聚类1 (Table 1)gydF4y2Ba1gydF4y2Ba;额外的文件gydF4y2Ba4gydF4y2Ba),与PPDK表达模式一致。作为淀粉合成的关键限速酶,AGPase催化的是一个完全可逆的反应,反应的方向取决于PPi和ATP的相对浓度。由此可见,ppdk主导的PEP对Pyr的形成可能会减少胞质PPi的积累,促进ADP-Glu的合成,从而促进淀粉的合成和积累。这一建议得到了另一项对水稻籽粒的研究结果的支持,该研究发现编码PPDK的基因突变表明了这种酶在淀粉合成中的作用的重要性[gydF4y2Ba72gydF4y2Ba].此外,玉米和水稻等物种的谷物胚乳是典型的缺氧组织,在籽粒发育过程中,内氧浓度的降低会抑制ATP的产生[gydF4y2Ba60gydF4y2Ba,gydF4y2Ba73gydF4y2Ba].因此,ppdk主导的PEP转化为Pyr的形成可能通过将AMP转化为ATP,并通过产生Pyr作为活性酒精发酵途径的底物来促进能量供应(见上文)。后一种途径在不消耗氧气的情况下产生ATP [gydF4y2Ba74gydF4y2Ba],并有助于在低氧张力下维持淀粉合成所需的适当ATP水平。PPDK在水稻根部低氧胁迫下的表达增强为其功能提供了进一步的线索[gydF4y2Ba75gydF4y2Ba].这些证据表明,PPDK可能优先作用于PEP向Pyr的形成方向,从而减少胞质PPi的积累,增加ATP含量,最终促进籽粒灌浆期淀粉合成和能量供应。gydF4y2Ba
14-3-3蛋白可能在籽粒发育过程中起重要作用gydF4y2Ba
在本研究中,14-3-3蛋白的三种亚型(B4FRG1、B6SZB9和B6T7L9)被鉴定为DEPs。有趣的是,这3种物质在早期(3-10 DAP)均表现出显著积累,然后逐渐降低到较低水平,直到籽粒成熟(c1;表格gydF4y2Ba1gydF4y2Ba).类似的表达模式也在其他物种中观察到,如蓖麻[gydF4y2Ba22gydF4y2Ba和大米[gydF4y2Ba15gydF4y2Ba这表明它可能在粮食发展中发挥作用。多项研究报道14-3-3蛋白参与多种细胞生理过程,如细胞信号转导、细胞周期调控、氮碳同化、防御机制[gydF4y2Ba76gydF4y2Ba,gydF4y2Ba77gydF4y2Ba].在大麦和玉米籽粒中,14-3-3蛋白可能与淀粉合成相关酶如ADPase、GBSS、SBE和ISA相互作用[gydF4y2Ba78gydF4y2Ba,gydF4y2Ba79gydF4y2Ba,这表明它们可能参与了淀粉合成的调控。有趣的是,在gydF4y2Ba拟南芥gydF4y2Ba叶片中14-3-3蛋白的减少或过表达分别与淀粉含量的急剧增加或减少相关[gydF4y2Ba80gydF4y2Ba,gydF4y2Ba81gydF4y2Ba].此外,对水稻籽粒的蛋白质组学和western blotting分析表明,14-3-3蛋白在高淀粉粒中的表达水平低于低淀粉粒[gydF4y2Ba82gydF4y2Ba].因此,14-3-3蛋白的高表达可能会降低淀粉合成相关酶的活性,从而不利于淀粉的形成和积累[gydF4y2Ba80gydF4y2Ba,gydF4y2Ba82gydF4y2Ba].与这一假设相一致的是,我们在这里发现14-3-3蛋白的最高表达水平发生在早期,而在10 DAP后淀粉合成酶开始上调,14-3-3蛋白的表达水平急剧下降。然而,14-3-3蛋白调控淀粉合成的潜在机制仍不清楚;确定这些蛋白质的作用可能对提高作物的淀粉产量有价值。gydF4y2Ba
结论gydF4y2Ba
研究了玉米籽粒3 - 50 DAP 8个连续发育阶段蛋白质表达的动态变化。应用iTRAQ技术鉴定了4751个蛋白质,鉴定出1235个蛋白质为籽粒发育过程中的DEPs,表明基于iTRAQ的定量蛋白质组分析是描述复杂代谢过程的有力技术。结果表明,籽粒不同发育阶段的代谢和细胞过程的协调性不同;例如,参与细胞生长/分裂的DEPs在早期后下调,而参与淀粉生物合成和防御/胁迫的DEPs在中后期分别显著上调。我们还证明了在不同发育阶段抗氧化系统中多种蛋白质的协调,这对维持ROS稳态至关重要。此外,一些DEPs,如锌金属硫蛋白II类、PPDK和14-3-3蛋白,在特定的发育阶段表达发生重大变化,表明它们在玉米籽粒发育中起着重要作用。这些结果为进一步了解影响玉米产量和品质的分子机制提供了新的线索。gydF4y2Ba
方法gydF4y2Ba
植物材料及取样gydF4y2Ba
本研究以中国玉米优良品种登海661 (DH351/DH372)为材料。该种子来自山东登海种子有限公司(中国莱州)。在山东农业大学泰安(东经36°10,北纬117°04)试验农场进行玉米生长季种植。对当天开花的植物进行标记,并进行人工自花授粉。3、5、10、15、20、30、40、50 DAP,每个阶段采集9穗。为了增加材料的均匀性,从每个穗的中间部分采样受精卵。在每个阶段,通过混合三个穗轴上相等数量的颗粒来制备三个样品;样品立即保存在−80°C,直到提取蛋白质。测定各粒期鲜重和干重。收集10-50 DAP和3-30 DAP的谷粒,分别测定淀粉总含量和胚乳细胞数量,如前所述[gydF4y2Ba83gydF4y2Ba].gydF4y2Ba
蛋白质的提取gydF4y2Ba
用研钵和研杵将谷物样品在液氮中磨成细粉末;粉末悬浮在10倍体积的预冷丙酮(−20°C)中,其中含有10% (v/v)三氯乙酸(TCA)。搅拌均匀后,在−20℃下沉淀2h。然后将匀浆在20000℃下离心30分钟gydF4y2BaggydF4y2Ba4°C,小心取出上清液;用冷丙酮冲洗三次,在−20°C放置30分钟,然后在20,000℃离心gydF4y2BaggydF4y2Ba在4°C下放置30分钟。将得到的微球溶解在含有8 M尿素、30 mM HEPES、1 mM聚乙烯聚吡烷酮(PMSF)、2 mM EDTA和10 mM二硫苏糖醇(DTT)的裂解缓冲液中,然后超声5分钟。将溶解的蛋白提取物在20000处离心gydF4y2BaggydF4y2Ba在4℃下30分钟,收集上清液,在56℃下用10 mM DTT还原1小时,然后与55 mM碘乙酰胺(IAM)在黑暗中烷基化1小时。使用5倍体积的冷丙酮在−20℃下沉淀3小时,然后在20000℃离心gydF4y2BaggydF4y2Ba将得到的颗粒溶于0.5 M三乙基碳酸氢铵(TEAB)缓冲液中,加入0.1% SDS,超声5 min,离心20,000℃gydF4y2BaggydF4y2Ba取上清液进行液体消化,以牛血清白蛋白(BSA)为标准,Bradford assay (Bio-Rad, Hercules, CA, USA)测定蛋白浓度。gydF4y2Ba
在溶液消化和iTRAQ标记gydF4y2Ba
每个样品在TEAB缓冲液中加入3.3 μL胰蛋白酶(1 μg/μL) (Promega, Madison, WI, USA),在100 μg蛋白质中,37℃消化24 h。加入新鲜胰蛋白酶(1 μL),再次消化12 h。沉淀在30 μL 0.5 M TEAB中溶解,与70 μL异丙醇混合。然后,根据制造商的说明,用iTRAQ试剂(AB SCIEX, Framingham, MA, USA)对消化后的肽进行标记。分别用iTRAQ试剂113、114、115、116、117、118、119和121对3、5、10、15、20、30、40和50 DAP样品进行标记。进行了三个独立的生物学实验。gydF4y2Ba
SCX和质/女士gydF4y2Ba
汇集的肽溶解在强阳离子交换(SCX)缓冲液A (10 mM磷酸氢钾单碱基(KHgydF4y2Ba2gydF4y2Ba阿宝gydF4y2Ba4gydF4y2Ba)在25%的乙腈中,pH值2.8)。用磷酸将混合物调整至pH值3,然后使用配备硅基SCX柱(250 × 4.6 mm, 5 μm, 100 Å, Phenomenex, Torrance, CA, USA)的高效液相色谱(HPLC)系统(日本岛adzu,京都,日本)进行分馏。在缓冲液B (10 mM KH)的流速为1 mL/min下,收集36个组分gydF4y2Ba2gydF4y2Ba阿宝gydF4y2Ba4gydF4y2Ba和2 M氯化钾(KCl)在25%的乙腈中,pH为2.8),梯度如下:0 % 45分钟,0 - 5% 1分钟,5 - 30% 20分钟,30 - 50% 5分钟和保持5分钟,50 - 100% 5分钟和保持10分钟。按照制造商的说明,馏分用层- x 33 μm PolyRevStage SPE (Phenomenex)脱盐,在离心速度真空浓缩器中冻干。然后,在每个干燥的分数管中加入0.1%甲酸30 μL,再溶解溶液0.1 μL在Anchor-chip平板的目标孔上进行MALDI-TOF检测。经MALDI-TOF (Bruker Daltonics, Germany)检验后,将36个馏分按峰面积合并为16个最终馏分。gydF4y2Ba
质谱分析使用Dionex Ultimate 3000 Nano LC系统连接到Q-Exactive质谱仪(Thermo Fisher Scientific, MA, USA)进行。采用Acclaim PePmap C18反相色谱柱(75 μm × 2 cm, 3 μm, 100 Å, Thermo Scientific)对多肽混合物进行分离,用反相C18色谱柱(75 μm × 10 cm, 5 μm, 300 Å, Agela Technologies)进行分离,梯度为5 - 80% (v/v)乙腈,0.1%甲酸,流速为300 nL/ min,溶剂a为0.1%甲酸水溶液,45 min。采用正离子模式进行全质谱(MS)扫描(350-2000 m/z),分辨率为70000 (200 m/z), AGC目标值为3-6,最大离子积累时间为50 MS,扫描次数范围为1次,动态排除时间为15 s。利用以下条件获得多肽和肽段m/z的信息:每次全扫描(MS2扫描)后收集20个片段文件,更高的碰撞能量分解(HCD)碎片,2 m/z的隔离窗口,全扫描分辨率为17500 (200 m/z),微扫描1次,最大离子积累时间为100 ms,标准化碰撞能量为28 eV,欠填充率为1%。gydF4y2Ba
数据分析gydF4y2Ba
蛋白质鉴定用Proteome Discoverer 1.3 (Thermo Fisher Scientific)处理MS原始文件,用内部吉祥物软件2.3.01 (Matrix Science, London, UK)进行搜索。用UniProt-自动搜索获得的MS/MS光谱gydF4y2BaZeamaysgydF4y2Ba蛋白质数据库(86,922条序列,2014年12月)。搜索参数如下:选择胰蛋白酶为允许漏切一次的酶;半胱氨酸氨基甲基化固定修饰;设置N端iTRAQ 8-plex修饰、K和Y、N端Gln→Pyro-Glu和蛋氨酸氧化为变量修饰;肽耐受设定为15 ppm;MS/MS公差设定为20 mmu。蛋白质鉴定和定量数据分析需要至少1个假发现率(FDR)≤1%的肽段。gydF4y2Ba
用两个标准对鉴定的蛋白质进行定量:1)选择中位蛋白质比;2)最小前体电荷设置为2,只使用唯一的多肽进行定量。在3 DAP下获得的标记样品作为参考(REF),基于每个识别肽的报告离子强度的加权平均。为了表明每个阶段蛋白质的丰度,计算每个阶段样品相对于3 DAP的相对蛋白质比,作为指定样品所有肽段的中位数(3 d/REF、5 d/REF、10 d/REF、15 d/REF、20 d/REF、30 d/REF、40 d/REF和50 d/REF)。在分析籽粒发育过程中DEPs时,只使用至少2个生物重复的定量信息。用3个生物重复的平均值表示每个阶段的最终蛋白丰度,定义在不同阶段平均蛋白丰度变化大于1.5倍(ANOVA检验,p≤0.05)的蛋白为DEPs。利用Cluster 3.0软件(gydF4y2Bahttp://bonsai.hgc.jp/~mdehoon/software/cluster/software.htmgydF4y2Ba),利用相似性度量和欧氏距离。设置集群数量为5个,使用关联的Java TreeView 1.1.1软件将结果可视化。gydF4y2Ba
缩写gydF4y2Ba
- 1 cys-prx:gydF4y2Ba
-
1-cys-peroxiredoxingydF4y2Ba
- 二:gydF4y2Ba
-
二维电泳gydF4y2Ba
- 6 pgdh:gydF4y2Ba
-
6-phosphogluconate脱氢酶gydF4y2Ba
- ADP-Glu:gydF4y2Ba
-
ADP-glucosegydF4y2Ba
- AGPase:gydF4y2Ba
-
ADP-glucose pyrophosophorylasegydF4y2Ba
- BT1:gydF4y2Ba
-
ADP-glucose brittle-1运输车gydF4y2Ba
- 衣冠楚楚的:gydF4y2Ba
-
授粉后一天gydF4y2Ba
- DEPs:gydF4y2Ba
-
差异表达的蛋白质gydF4y2Ba
- G1P:gydF4y2Ba
-
glucose-1-phosphategydF4y2Ba
- G6PDH:gydF4y2Ba
-
glucose-6-phosphate 1-dehydrogenasegydF4y2Ba
- gbs:gydF4y2Ba
-
granule-bound淀粉合酶gydF4y2Ba
- ISA:gydF4y2Ba
-
异淀粉酶gydF4y2Ba
- iTRAQ:gydF4y2Ba
-
等压标记用于相对和绝对定量gydF4y2Ba
- 李:gydF4y2Ba
-
胚胎晚期丰富gydF4y2Ba
- 动员:gydF4y2Ba
-
磷酸烯醇丙酮酸gydF4y2Ba
- PPDK:gydF4y2Ba
-
丙酮酸正磷酸盐dikinasegydF4y2Ba
- 购买力平价:gydF4y2Ba
-
磷酸戊糖途径gydF4y2Ba
- Pyr:gydF4y2Ba
-
丙酮酸gydF4y2Ba
- RNA-seq:gydF4y2Ba
-
RNA序列gydF4y2Ba
- ROS:gydF4y2Ba
-
活性氧gydF4y2Ba
- SBE:gydF4y2Ba
-
淀粉分支酶gydF4y2Ba
- 瑞士:gydF4y2Ba
-
可溶性淀粉合酶gydF4y2Ba
- 苏西:gydF4y2Ba
-
蔗糖合酶gydF4y2Ba
- 柠檬酸:gydF4y2Ba
-
三羧酸gydF4y2Ba
参考文献gydF4y2Ba
Prioul JL, Méchin V, Lessard P, Thévenot C, Grimmer M, Chateau-Joubert S,等。玉米胚乳发育和淀粉填充的转录组、蛋白质组和代谢分析。植物学报;2008;6:855 - 863。gydF4y2Ba
奥尔森OA。胚乳发育。细胞化和细胞命运说明。植物学报2001;20(3):393 - 393。gydF4y2Ba
Sabelli PA, Larkins BA。胚乳:禾本科植物胚乳的发育植物杂志。2009;149:14-26。gydF4y2Ba
张志强,张志强。玉米胚乳中淀粉合成的研究。《植物学报》1995;46:465 - 468。gydF4y2Ba
刘志强,刘志强,刘志强,刘志强,刘志强。小麦胚乳中淀粉生物合成的研究进展。植物生理化学。2010;48:383 - 392。gydF4y2Ba
James MG, Denyer K, Myers AM。谷物胚乳中淀粉的合成。植物学报。2003;6:215 - 222。gydF4y2Ba
金旭,傅智,丁丹,李伟,刘智,唐军。玉米籽粒灌浆率相关基因的蛋白质组学鉴定。《公共科学图书馆•综合》。2013;8:e59353。gydF4y2Ba
Méchin V, Thévenot C, Le Guilloux M, Prioul JL, Damerval C.玉米胚乳蛋白质组发育分析表明丙酮酸正磷酸二激酶起着关键作用。植物杂志。2007;143:1203-19。gydF4y2Ba
李光,王东,杨锐,Logan K,陈宏,张帅,等。通过转录组测序鉴定玉米胚乳发育过程中基因表达的时间模式。中国科学(d辑)2014;gydF4y2Ba
陈杰,曾斌,张明,谢松,王刚,Hauck A,等。玉米胚和胚乳发育的动态转录组景观。植物杂志。2014;166:252 - 64。gydF4y2Ba
Verza NC, e Silva TR, Neto GC, Nogueira FT, Fisch PH, de Rosa Jr VE等。通过对表达序列标签的全转录组分析,揭示了玉米基因的胚乳偏好表达。植物化学。2005;29(3):393 - 393。gydF4y2Ba
刘志强,刘志强,刘志强,等。玉米发育过程中的转录全基因组图谱。植物j . 2011; 66:553 - 63。gydF4y2Ba
张志强,张志强,张志强,张志强等。玉米胚成熟的转录组分析。植物生物学。2013;13:19。gydF4y2Ba
王伟,沈智,吴克杰,吴克斌,张志刚,等。玉米种子蛋白质型图谱的蛋白质网络重建。中国地质大学学报(自然科学版)2013;gydF4y2Ba
徐世斌,李涛,邓志勇,冲凯,薛勇,王涛。蛋白质组学动态分析揭示水稻灌浆籽粒中心碳代谢与酒精发酵之间的切换。植物杂志。2008;148:908-25。gydF4y2Ba
胡刚,高杰,刘俊,K集团,陈森,温德尔。野生和驯化棉纤维发育的蛋白质组学分析gydF4y2Ba海岛棉gydF4y2Ba.新植醇。2013;200:570 - 82。gydF4y2Ba
刘志强,刘志强,刘志刚,等。拟南芥种子灌浆的系统分析:使用一般线性模型评估转录本和蛋白表达的一致性。植物杂志。2010;152:2078 - 87。gydF4y2Ba
Tasleem-Tahir A, Nadaud I, Girousse C, Martre P, Marion D, Branlard G.小麦胚层蛋白质组学分析(gydF4y2Ba小麦gydF4y2Bal .)粮食发展。蛋白质组学。2011;11:371-9。gydF4y2Ba
徐慧,张伟,高艳,赵艳,郭玲,王杰。水稻胚胎发育的蛋白质组学分析(英文)gydF4y2Ba栽培稻gydF4y2Ba).足底。2012;235:687 - 701。gydF4y2Ba
杜邦调频。小麦的代谢途径(gydF4y2Ba小麦gydF4y2Ba)蛋白质组学显示胚乳淀粉质体。植物生物学。2008;8:39。gydF4y2Ba
李志强,李志强,李志强,等。大麦籽粒灌浆与成熟的蛋白质组学分析。植物杂志。2002;129:1308-19。gydF4y2Ba
休斯顿NL, Hajduch M, Thelen JJ。蓖麻种子灌浆的定量蛋白质组学研究:与大豆和油菜籽比较,揭示了蓖麻种子光合代谢与非光合代谢的差异。植物杂志。2009;151:857 - 68。gydF4y2Ba
李志强,李志强,李志强gydF4y2BaMedicago truncatulagydF4y2Ba种子发育建立了与储量积累相关的各种代谢过程的时间框架。植物杂志。2003;133:664 - 82。gydF4y2Ba
Hajduch M, Ganapathy A, Stein JW, Thelen JJ。大豆籽粒灌浆系统蛋白质组学研究。建立高分辨率的二维参考图、表达谱和交互式蛋白质组数据库。植物杂志。2005;137:1397 - 419。gydF4y2Ba
Agrawal GK, Hajduch M, Graham K, Thelen JJ。大豆种子灌浆蛋白质组的深入研究和与油菜籽的平行研究的比较。植物杂志。2008;148:504-18。gydF4y2Ba
张志强,张志强,张志强,等。双向凝胶电泳:样品制备、检测和定量的最新进展。中国生物医学杂志2002;6:46-50。gydF4y2Ba
吴文武,王贵,白世杰,沈瑞夫。采用2D凝胶或LC-MALDI TOF/TOF对DIGE、cICAT和iTRAQ三种蛋白质组学定量方法进行比较研究。中国生物医学工程学报2006;5:651 - 651。gydF4y2Ba
Karp NA, Huber W, Sadowski PG, Charles PD, Hseter SV, Lilley KS。解决iTRAQ定量中的准确性和精度问题。细胞蛋白质组学。2010;9:1885-97。gydF4y2Ba
质谱技术在蛋白质组学中的应用。植物学报,2010;31(5):393 - 393。gydF4y2Ba
子杰,张杰,王强,周斌,钟杰,张超,等。应激反应蛋白在水稻(gydF4y2Ba栽培稻gydF4y2Ba定量蛋白质组学分析表明,胚胎发生。《公共科学图书馆•综合》。2013;8:e74229。gydF4y2Ba
马超,周军,陈光,边勇,吕丹,李欣,等。基于itraq的定量蛋白质组学和磷酸化分析揭示了小麦籽粒发育过程中中枢代谢的变化。BMC基因组学。2014;15:1029。gydF4y2Ba
Méchin V, Balliau T, Chateau-Joubert S, Davanture M, Langella O, Negroni L等。玉米胚乳蛋白质组的二维图谱。植物化学。2004;65:1609-18。gydF4y2Ba
辛欣,林晓华,周永昌,陈晓林,刘旭,吕XX。玉米种子蛋白质组分析:人工老化的影响。杂志。2011;143:126-38。gydF4y2Ba
黄H, Møller IM,宋sq。玉米胚发育和萌发过程中脱水耐性的蛋白质组学研究。蛋白质组学。2012;75:1247 - 62。gydF4y2Ba
吴旭,刘慧,王伟,陈松,胡旭,李超。玉米种子活力的蛋白质组学分析。植物学报。2011;33:181-91。gydF4y2Ba
微管细胞骨架研究。植物科学。2002;5:494-501。gydF4y2Ba
Herrera I, De La Paz Sánchez M, Molina J, Plasencia J, Vázquez-Ramos JM。玉米籽粒发育和萌发中增殖细胞核抗原的表达:植物激素的调控及其与假定的细胞周期蛋白的关联杂志。2000;110:127-34。gydF4y2Ba
王志强,王志强,王志强,等。MAIZEWALL。玉米细胞壁生物合成和组装的数据库和发育基因表达谱。植物杂志。2007;143:339 - 63。gydF4y2Ba
Martin SW, Glover BJ, Davies JM。脂质微域-植物膜得到组织。植物科学进展。2005;10:335 - 335。gydF4y2Ba
一个简短的导游,通过功能和结构特征的皂苷样蛋白。j . 2005; 389:249-57。gydF4y2Ba
Cochrane CG, Revak SD。肺表面活性蛋白B (SP-B):结构-功能关系。科学。1991;254:566-8。gydF4y2Ba
刘文华,李文华,李文华。泛素-蛋白酶体途径与植物发育的关系。植物细胞。2004;16:3181 - 95。gydF4y2Ba
与植物生物学联系的泛素- 26s蛋白酶体系统。中国生物医学杂志。2009;10:385 - 397。gydF4y2Ba
黄晓明,刘文华。磷酸戊糖氧化途径:结构与组织。植物学报。2003;6:236-46。gydF4y2Ba
Horecker BL,戊糖磷酸途径。生物化学学报2002;277:47965-71。gydF4y2Ba
王芳,孙志强,刘志强,等。蔗糖合酶对番茄果实淀粉积累的影响。植物杂志。1993;101:321-7。gydF4y2Ba
刘坤昌,刘庆昌,刘庆昌,刘庆昌。糖腺苷酸转运蛋白britte -1在玉米胚乳淀粉质体中表达的研究进展。植物杂志。1998;117:1235-52。gydF4y2Ba
Bowsher CG, scrasfield EF, Esposito S, Emes MJ, Tetlow IJ。adp -葡萄糖在谷物胚乳淀粉质包膜上的转运特性。J Exp Bot. 2007; 58:1321-32。gydF4y2Ba
李志强,李志强,李志强,等gydF4y2Ba蜡状gydF4y2Ba在玉米轨迹。细胞。1983;35:225-33。gydF4y2Ba
Cao H, Imparl-Radosevich J, Guan H, Keeling PL, James MG, Myers AM。玉米胚乳可溶性淀粉合酶活性的鉴定。植物杂志。1999;120:205-16。gydF4y2Ba
姚明Y,汤普森DB,吉尔蒂南MJ。玉米淀粉分枝酶异构体和支链淀粉结构。在缺乏淀粉分支酶IIb的情况下,淀粉分支酶Ia的进一步缺乏导致分支增加。植物杂志。2004;136:3515-23。gydF4y2Ba
黄建平,李建平,李建平,等。水稻胚乳中异淀粉酶和葡萄糖焦磷酸化酶在淀粉粒合成中的作用。植物j . 2005; 42:164 - 74。gydF4y2Ba
刘志强,刘志强,刘志强,刘志强,等。异质和同聚异淀粉酶型淀粉脱支酶在玉米胚乳发育中的作用。植物杂志。2010;153:956 - 69。gydF4y2Ba
张志刚,张志勇,张志勇,张志勇,等。水稻塑性α-葡聚糖磷酸化酶基因的突变影响了胚乳淀粉的合成和结构。植物细胞。2008;20:1833-49。gydF4y2Ba
黄sk, A Nishi, H佐藤,TW Okita。水稻胚乳特异可塑性α-葡聚糖磷酸化酶是合成短链麦芽低聚糖的重要酶类。中国科学:地球科学。2010;gydF4y2Ba
曾建华,黄阿。植物种子油体的表面结构与性质。中国生物医学杂志1992;117:327-35。gydF4y2Ba
Siloto RM, Findlay K, Lopez-Villalobos A, yeec, Nykiforuk CL, Moloney MM.油苷的积累决定了拟南芥种子油体的大小。植物细胞。2006;18:1961 - 74。gydF4y2Ba
张志强,张志强,张志强,张志强,等。玉米LEA蛋白的研究:从蛋白质组学到功能方法。植物学报。2012;53:312-29。gydF4y2Ba
Battaglia M, Olvera-Carrillo Y, Garciarrubio A, Campos F, Covarrubias AA。神秘的LEA蛋白和其他亲水性蛋白。植物杂志。2008;148:6-24。gydF4y2Ba
刘志强,刘志强,刘志强,等。玉米籽粒淀粉/脂质平衡及其资源分配的位置线索。植物j . 2005; 42:69 - 83。gydF4y2Ba
李志强,李志强,李志强,等。植物活性氧基因网络研究进展。中国植物科学。2004;gydF4y2Ba
氧化还原稳态和抗氧化信号:应激知觉和生理反应之间的代谢接口。植物细胞。2005;17:1866 - 75。gydF4y2Ba
Stacy RA, Nordeng TW, Culianez-Marcia FA, aallen RB。与休眠相关的过氧化物还蛋白抗氧化剂PER1定位于大麦胚核和糊粉层细胞。植物j . 1999; 19:1-8。gydF4y2Ba
李高,张德宏,郑bg,池耀辉,李志勇,崔洋,等。水稻cys -过氧化物还蛋白在转基因烟草中过表达后不保持休眠状态,反而增强抗氧化活性。2月。2000;486:103-6。gydF4y2Ba
Haslekås C, Viken MK, Grini PE, Nygaard V, Nordgard SH, Meza TJ,等。种子1-半胱氨酸过氧化物还蛋白抗氧化剂不参与休眠,但有助于抑制萌发期间的胁迫。植物杂志。2003;133:1148-57。gydF4y2Ba
Kim SY, Paeng SK, Nawkar GM, Maibam P, Lee ES, Kim KS,等。1-Cys过氧化物还蛋白是种子休眠的调节因子,在氧化应激条件下起着分子伴侣的作用。植物科学。2011;181:119-24。gydF4y2Ba
刘建军,刘建军,刘建军。细胞内金属硫蛋白对镉毒性的保护作用。《药理学进展》1999;39:267-94。gydF4y2Ba
郭文杰,Bundithya W, Goldsbrough PB拟南芥金属硫蛋白基因家族的特征:衰老和铜胁迫下组织特异性表达和诱导。新植醇。2003;159:369 - 81。gydF4y2Ba
川岛I, Kennedy TD, Chino M, Lane BG。小麦Ec金属硫蛋白基因:像哺乳动物的锌gydF4y2Ba2 +gydF4y2Ba金属硫蛋白基因在胚胎发生过程中显著表达。欧洲生物化学杂志1992;209:971-6。gydF4y2Ba
玉米丙酮酸、正磷酸盐二激酶基因差异表达的分子机制。植物细胞。1991;3:225-45。gydF4y2Ba
Chastain CJ, Heck JW, Colquhoum TA, Voge DG, Gu XY。丙酮酸磷酸二激酶在水稻发育中的翻译后调控(gydF4y2Ba栽培稻gydF4y2Ba)种子。足底。2006;224:924-34。gydF4y2Ba
Kang HG, Park S, Matsuoka M, An G.水稻白核胚乳粉胚乳-4由c型丙酮酸正磷酸盐二激酶基因敲除突变产生(gydF4y2BaOsPPDKBgydF4y2Ba).植物j . 2005; 42:901-11。gydF4y2Ba
许世斌,于海涛,闫丽芬,王涛。水稻贮藏期胚乳蛋白质组学与细胞学研究。中国生物医学工程学报2010;9:4906-18。gydF4y2Ba
植物代谢对缺氧的反应。植物学报。2003;6:247-56。gydF4y2Ba
低氧胁迫和水分亏缺诱导C3植物水稻根部胞质丙酮酸磷酸二激酶(PPDK)表达。植物j . 1998; 15:89 - 98。gydF4y2Ba
骑士D,莫里斯ER,沃克JC。14-3-3和FHA结构域介导磷酸化蛋白相互作用。植物学报。2009;60:67-91。gydF4y2Ba
植物14-3-3蛋白赶上了它们的哺乳动物同源蛋白。植物学报。2009;12:60 - 5。gydF4y2Ba
Alexander RD, Morris PC。大麦籽粒14-3-3结合蛋白的蛋白质组学分析。蛋白质组学。2006;6:1886 - 96。gydF4y2Ba
窦勇,刘旭,尹勇,韩松,陆勇,刘勇,等。亲和层析揭示了两个14-3-3蛋白在玉米籽粒发育中的独特功能。蛋白质组学。2015;114:274 - 86。gydF4y2Ba
申克PC,钟焕杰,吴凯,费尔RJ。颗粒相关植物14-3-3蛋白对淀粉积累的调控中国科学(d辑)2001;98:765-70。gydF4y2Ba
Diaz C, Kusano M, Sulpice R, Araki M, Redestig H, Saito K,等。确定拟南芥14-3-3蛋白在中枢代谢过程中的新功能。BMC生物学报。2011;5:192。gydF4y2Ba
张智,赵华,唐杰,李智,李智,陈丹,等。水稻灌浆不良分子机制的蛋白质组学研究(gydF4y2Ba栽培稻gydF4y2Bal .)下的小穗。《公共科学图书馆•综合》。2014;9:e89140。gydF4y2Ba
刘晔,刘鹏,董st,张景文。正常玉米与高油玉米杂交籽粒灌浆过程中色散效应引起的激素变化作物科学。2010;50:215-21。gydF4y2Ba
确认gydF4y2Ba
基金资助:国家自然科学基金项目(31371576),国家重点研发计划项目(2016YFD0300106;国家科技支撑计划项目(2013BAD07B06-2),国家公益性产业(农业)计划(201203100;201203096),山东省现代农业技术产业体系(SDAIT-02-08),山东省农业科技创新项目,山东省玉米育种与栽培技术重点实验室,山东省高等学校科技计划项目(J14LF10)。gydF4y2Ba
资金gydF4y2Ba
不适用。gydF4y2Ba
数据和材料的可用性gydF4y2Ba
支持本文结论的数据集包含在本文及其附加文件中。gydF4y2Ba
作者的贡献gydF4y2Ba
YT和LG进行了所有的实验和数据分析。ZJ和ZB进行了现场试验和取样。DS和LP构思了研究,策划了实验,并帮助起草了稿件。所有作者阅读并批准最终稿。gydF4y2Ba
作者的信息gydF4y2Ba
不适用。gydF4y2Ba
相互竞争的利益gydF4y2Ba
作者声明他们之间没有利益冲突。gydF4y2Ba
同意出版gydF4y2Ba
不适用。gydF4y2Ba
伦理认可和同意参与gydF4y2Ba
不适用。gydF4y2Ba
作者信息gydF4y2Ba
从属关系gydF4y2Ba
相应的作者gydF4y2Ba
附加文件gydF4y2Ba
额外的文件1:gydF4y2Ba
玉米籽粒中蛋白质含量(XLSX 745 kb)gydF4y2Ba
额外的文件2:gydF4y2Ba
定量蛋白表达谱数据(XLSX 1136kb)gydF4y2Ba
额外的文件3:gydF4y2Ba
玉米籽粒中未鉴定蛋白的同源性(XLSX 48 kb)gydF4y2Ba
额外的文件4:gydF4y2Ba
DEPs被分配给功能类别和集群成员(XLSX 166kb)gydF4y2Ba
权利和权限gydF4y2Ba
开放获取gydF4y2Ba本文根据知识共享署名4.0国际许可协议(gydF4y2Bahttp://creativecommons.org/licenses/by/4.0/gydF4y2Ba),它允许不受限制地在任何媒体上使用、分发和复制,前提是您要适当地注明原作者和来源,提供创作共用许可的链接,并说明是否进行了更改。知识共享公共领域转让豁免书(gydF4y2Bahttp://creativecommons.org/publicdomain/zero/1.0/gydF4y2Ba)适用于本文提供的数据,除非另有说明。gydF4y2Ba
关于这篇文章gydF4y2Ba

引用这篇文章gydF4y2Ba
于涛,李国强,董硕。gydF4y2Baet al。gydF4y2Ba利用iTRAQ对玉米籽粒发育进行蛋白质组学分析,揭示了不同代谢过程的时间程序。gydF4y2BaBMC植物杂志gydF4y2Ba16日,gydF4y2Ba241(2016)。https://doi.org/10.1186/s12870-016-0878-1gydF4y2Ba
收到了gydF4y2Ba:gydF4y2Ba
接受gydF4y2Ba:gydF4y2Ba
发表gydF4y2Ba:gydF4y2Ba
DOIgydF4y2Ba:gydF4y2Bahttps://doi.org/10.1186/s12870-016-0878-1gydF4y2Ba
关键字gydF4y2Ba
- 玉米gydF4y2Ba
- 粮食发展gydF4y2Ba
- 蛋白质组学gydF4y2Ba
- iTRAQgydF4y2Ba
- 淀粉gydF4y2Ba